
پیشرفتهای جدید در درک مکانیسمهای بیماریزایی قارچهای فرصت طلب
قسمت نخست
دكتراي علوم آزمايشگاهي، قارچ شناسي پزشكي دکتر محمد قهری
آزمایشگاه تشخیص طبی رسالت

در ابتدا لازم میدانم که از زحمات آقایان احمدرضا براهیمی و سعید پرهام دانشجویان پرتلاش دوره کارشناسی ارشد قارچ شناسی پزشکی دانشگاه تربیت مدرس که در مراحل نخست ترجمهی این متن با اینجانب همکاری کردند صمیمانه تشکر و قدردانی نمایم.
مقدمه:
پاتوژنهای قارچی
قارچها اجزاء مهمی از اکوسیستم هستند؛ چرخه جهانی کربن را هدایت میکنند و همچنین از طریق جمعیتهای مایکوریزی در خاک سبب بقای کشاورزی و تنوع گیاهی میشوند، همچنین از طرق دیگری همانند تأمین مواد غذایی، تولید مشروبات الکلی و آنتیبیوتیکهای حافظ حیات، در خدمت بشر هستند.
اولین شواهد فسیل قارچی (آربسکولار مایکوریزآه) که 460 میلیون سال عمر دارد به طور مستقیم مرتبط با گیاهان است. قارچها در برخی از مراحل سیر تکاملی خود این توانایی را به دست میآورند که به صورت پاتوژن تبدیل شوند و یا از موجودات زنده به عنوان منابع مغذی خود استفاده کنند تا چرخه زندگی خود را تکمیل کنند. آنالیزهای فیلوژنتیک اخیر تغییر و تحولات متعددی را بین سبکهای زندگی قارچهای پاتوژنیک و ساپروفیتیک نشان داده است. واضح است که توسعه متناظر با آن، مقاومت در برابر پاتوژنهای قارچی را ایجاد میکند؛ بدین صورت که استرینهائی از پاتوژن قارچی که فاقد ژن غالب Avrبود میتوانست سبب ایجاد بیماری شود تا زمانی که یک جهش اتفاقی ژن غالب دیگر (ژن R) که سبب بروز مقاومت بود را به جمعیت گیاه میآورد که منجر به ایجاد مقاومت میشد. در طول زمان یک مسابقهی تسلیحاتی واقع میشد؛ جائی که یک جهش تصادفی در جمعیت قارچها منجر به پیدایش یک ژن جدید Avr غیر قابل تشخیص در جمعیت گیاه میگردید که سبب ایجاد بیماری در گیاه میشد و این بیماری تا زمانی باقی میماند که یک ژن R دیگر در جمعیت گیاهی ظاهر شود و شرایط مقاومت به بیماری را مهیا کند. اگرچه ثابت شده است که این مدل بسیار پیچیدهتر از این واکنشهای ژنی تک محصولی میباشد، امروزه به خوبی مشخص شده است که پاتوژنهای قارچی گیاهی مسیر تکاملی خوبی را در ارتباط با اکتساب مکانیسمهای پیچیدهی تهاجم، طفره رفتن و فائق آمدن بر پاسخهای دفاعی گیاه پیمودهاند. در مقایسه با عفونتهای قارچی گیاهی، عفونتهای قارچی که پستانداران را درگیر میکنند نسبتاً نادر هستند. این امر قویاً بیان میکند که پستانداران مکانیسمهای دفاعی پیچیدهتری علیه قارچها را کسب کردهاند و مهم اینکه قارچها قادر نیستند مکانیسمهای بیماریزایی را در مقابل مقاومت پستانداران گسترش دهند و بعبارت دیگر مقاومت پستانداران را بیاثر سازند. اخیراً این فرضیه ارائه شده است که ظهور پستانداران در اواخر دورهی کرتاسه با توانایی آنها در زنده ماندن در مقابله با چالشهای عظیم مربوط به جمعیتهای قارچی که در این دوره رخ داده است در ارتباط است. علیرغم تماس روزانهی پستانداران با کنیدیها و هایفهای قارچی، به طور مشخصی پستانداران نسبت به قارچها مقاوم هستند. فهمیدن این مکانیسم مقاومت پستانداران و اینکه چگونه آنها از بروز بیماریهای قارچی جلوگیری میکنند برای درک پاتوژنز قارچها در انسان ضروری است.
مکانیسمهای این مقاومت قابل توجه احتمالاً به سبب ویژگیهای ارثی متعدد پستانداران است؛ نخست اینکه دمای مرکزی بدن بسیاری از پستانداران بین 39-37 درجه سانتیگراد میباشد. اکثریت قریب به اتفاق قارچها در دمای بین 25 تا 35 درجه سانتیگراد رشد میکنند و بنابراین رشد درون بدن میزبان پستاندار برای بسیاری از قارچها مجاز و امکان پذیر نیست. دوم اینکه پستانداران مایعات بدنی قلیایی دارند و این در حالی است که بسیاری از قارچها ترجیح میدهند که در یک PH اسیدی ملایم تا مایل به خنثی رشد کنند. سوم آنکه پستانداران سیستمهای ایمنی ذاتی و اکتسابی پیچیدهای کسب کردهاند که از رشد قارچ در زمانی که کنیدیهای قارچ استنشاق میشود جلوگیری میکنند. ترکیب این سه مکانیسم سد ممنوعهای را برای عفونتهای قارچی در انسان ایجاد میکند. برای اینکه عفونتهای قارچی بتوانند در انسان رخ دهند عوامل قارچی باید بتوانند بر این سد فائق آیند.
در این نوشتار در نهایت با این موضوع سر و کار داریم که قارچها چگونه بر این سد فائق آمده و تا کلونیزه شدن و ایجاد بیماری در پستانداران پیش میروند.
وقایع اخیر در پزشکی جدید به طور چشمگیری روابط بین قارچها و پستانداران را تغییر داده است. پیشرفتهای حاصله در درمانهای پزشکی، پیوند عضو، عفونتهای HIV و افزایش جمعیت افراد سالمند همگی در دو دههی گذشته سبب افزایش عفونتهای قارچی تهدید کننده حیات شدهاند.
این بیماریهای زمینهای و همچنین تکنولوژیهای مربوطه سبب افزایش جمعیت افراد دارای نقص ایمنی شدهاند. این دسته از افراد نسبت به عوامل قارچی خاصی حساس هستند که میتوانند به فاکتورهای ذاتی از قبیل دما و نیز سدهای ساختاری و شیمیایی میزبانان پستاندار غلبه کرده و در نتیجه سبب ایجاد عفونت شوند. عفونتهای قارچی در افراد دارای نقص ایمنی با عنوان “فرصت طلب” نامگذاری میشوند زیرا قارچهایی که عموماً این عفونتها را سبب میشوند ارگانیسمهای ساپروفیتیک هستند که قابلیت ایجاد بیماری در افراد دارای سیستم ایمنی سالم را ندارند.
اگرچه اخیراً افزایش این عفونتهای قارچی مهلک انسانی منجر به افزایش تلاشهای همسو در جهت شناخت بهتر مکانیسمهای پاتوژنز در پستانداران شده است، تلاشهای اخیر تحقیقاتی در زمینهی مکانیسم پاتوژنز بیماریهای قارچی در پستانداران بیشتر بر روی این قضیه متمرکز است که آیا این پاتوژنهای فرصت طلب برای ایجاد بیماری، فاکتورهای ویرولانس خاصی را در طول تکامل خود مانند همتاهای پاتوژنیک گیاهی خود بدست آوردهاند و یا اینکه به آسانی با محیط میزبانی که از پاسخهای ایمنی ذاتی و اکتسابی تهی شده بخوبی سازگاری پیدا کردهاند.
پیشرفتهای تکنولوژیک اخیر در زمینههای زیست شناسی ملکولی و علوم مبتنی بر ژنومیک شروع به یافتن پاسخ به این سؤال مهم کردهاند. هدف این نوشتار مروری بر پیشرفتهای تازه در درک ما از این مطلب است که چگونه پاتوژنهای قارچی فرصت طلب مهم قادر به ایجاد بیماری در میزبانان پستاندار هستند و تأکید بر اینکه چگونه فهم بیولوژی این ارگانیسمها در روشن ساختن مکانیسمهای بیماریزائی قارچهای پاتوژن انسانی اهمیت دارد. اگرچه در دو دههی اخیر پیشرفتهای چشمگیری در زمینه پاتوژنز عوامل قارچی انجام گرفته اما این نوشته برای هر عامل قارچی که مورد بحث قرار میگیرد جامع نیست. در اینجا مواردی و آن هم در سطح ملکولی مثال زده و مورد بحث قرار خواهد گرفت تا نشان داده شود که قارچهای فرصت طلب چگونه باعث ایجاد بیماری و عفونت در افراد دارای نقص ایمنی میشوند. در انتها بعنوان نتیجه گیری به طور خلاصه به ملاحظات مهم مربوط به عفونت در میزبان پرداخته خواهد شد.
ویرولانس فاکتور چیست؟
قبل از بحث در مورد “مکانیسم پاتوژنز قارچی” باید معنای دو واژه ی بحث برانگیز ویرولانس و پاتوژنز را به خوبی دانست.
متون مربوط به پاتوژنز میکروبی پر از مباحث و منازعات مربوط به این لغات بوده و اغلب بد بکار برده شدهاند، بنابراین درک درست از این موضوع که این لغات به تنهایی چگونه استفاده میشوند در فهمیدن مکانیسم پاتوژنز قارچی مهم است. مباحثه کامل در مورد تاریخچه پیچیدهی این اصطلاحات مورد بحث ما نیست و کسانی که مایل هستند در این مورد اطلاع پیدا کنند به مقاله زیر که در مورد تاریخچهی اصطلاحات (ویرولانس) و (پاتوژنز) است ارجاع داده میشوند.
[Casadevall A. Pirofski I. Host-pathogen interactions: the attributes of virulence. J Infect Dis 184: 337, 2001]
در اینجا بطور خلاصه درباره این موضوع بحث میکنیم که چگونه استفاده صحیح از این اطلاعات در درک مکانیسم پاتوژنز قارچهای بیماریزای انسانی ضروری است.
استفاده گسترده و متنوع از لغات (pathogenesis)، (virulence) و (virulence factor) ارتباطات بین دانشمندان در حیطههای مختلف تحقیقات مربوط به پاتوژنز میکروبی را محدود کرده و مهمتر اینکه این قضیه مانع از درک صحیح بیولوژی و تاریخچهی تکاملی این پاتوژنهای مهم شده است.
از لحاظ تاریخی اصطلاحات (pathogenesis) و (virulence) بر اساس توانایی میکروبها در ایجاد بیماری تعریف شدهاند، در نتیجه استفاده از این اصطلاحات پاتوژن محور بوده و نشان میدهد که یک انتخاب اختصاصی که از طرف میزبانان فشار میآورد منجر به تکامل فاکتور ویرولانس میشود.
اصطلاح (pathogenesis) دارای ریشهی یونانی(phato) به معنی (بیماری) و (genesis) به معنی (پیشرفت یا توسعه) میباشد و بنابراین اصطلاح (pathogenesis) به سادگی بیان میدارد که پاتوژنز یعنی: توانایی یک ارگانیسم در ایجاد بیماری.
در مقابل واژهی (virulence) به صورت “توانایی نسبی یک پاتوژن برای ایجاد بیماری” معنی میشود.
بنابراین همهی پاتوژنهای قارچی ایجاد بیماری میکنند اما برخی بیماریزاتر از بقیه هستند؛ به عبارت دیگر آنها توانایی بیشتری در ایجاد بیماری و آسیب در میزبان دارند. البته مطمئناً این قضیه این سؤال را ایجاد میکند که چرا برخی از قارچهای پاتوژن نسبت به سایرین دارای قدرت بیماریزایی بیشتری هستند؟
پاسخ به این سؤال در اصطلاح بحث برانگیز (virulence factor) نهفته است. (virulence factor) به عنوان اجزایی از یک ارگانیسم پاتوژن که اجازه میدهند آن پاتوژن ایجاد بیماری کند تعریف شده است.
به طور معمول این اجزاء به صورت ژنتیکی در ژنوم پاتوژن رمزگذاری شدهاند. حذف این اجزاء از طریق القاء جهش در آزمایشگاه و یا حذف در طی فرآیند تکاملی در طبیعت، ویرولانس پاتوژن را کاهش میدهد ولی اثری روی قابلیت زیستن ارگانیسم ندارد. بدینسان، فاکتورهای ویرولانس عملکردهای خاصی را که برای بیماریزانی نیاز دارند در سیر تکاملی خویش کسب کردهاند. ما این جزء تکاملی در تعریف فاکتور ویرولانس را برای درک بهتر بیماریزایی میکروبی در نظر میگیریم. با به حساب آوردن تاریخچهی تکامل احتمالی یک ژن یا محصولات ژنی میتوان بین ویرولانس فاکتور حقیقی و فاکتور ویرولانسی که به سادگی فقط از این جهت که در صورت غیاب آن، ارگانیسم قادر به ایجاد بیماری نیست، تفاوت قائل شد؛ برای مثال ژنهایی که در شرايط آزمایشگاهی حضورشان برای رشد قارچی اساسی بوده است زمانی در قارچ شناسی پزشکی به عنوان ویرولانس فاکتور نامیده شده بودند. البته امروزه مشخص شده است که این ژنها نقش کمی در پاتوژنز میکروبی دارند در عوض این ژنها در عملکرد تکاملی خود در بیولوژی پایهی ارگانیسم قارچی نقش مؤثری ايفا ميكنند. برداشت این نوع از ژنها موجب جلوگیری از رشد و یا بروز تغییراتی در رشد و سازگاری قارچ در شرايط آزمایشگاهی میشود، اما به طور طبیعی این استرینهای قارچی قادر به ایجاد بیماری در شرایط درون موجود زنده نخواهند بود، بنابراین اگر ما این تعریف کلاسیک را برای ویرولانس و ویرولانس فاکتور بپذیریم، بسیاری از قارچهای پاتوژن انسانی دارای فاکتورهای ویرولانس کلاسیک نیستند. با این حال وقتی شخصی متون قارچ شناسی را در مورد بیماریزائی قارچهای انسانی بررسی میکند، معلوم کردن اینکه فاکتور ویرولانس چه چیزی هست یا نیست مشکل است. این سردرگمی از آنجا ايجاد ميشود که برای بسیاری از قارچهای پاتوژن انسانی یکی از شروط اصلی بیماریزایی وجود نقص در سیستم ایمنی میزبان میباشد؛ به عبارت دیگر بسیاری از قارچهای بیماریزا توانایی غلبه بر سیستم ایمنی سالم و دست نخورده را ندارند.
امروزه در مورد استفاده از واژه (virulence factor) در ارتباط با این ژنهای مهم در فیزیولوژی نرمال قارچی و توانائی میکروبی بحثهایی وجود دارد که اغلب گمراه کننده است و مانع از درک بیولوژی این ارگانیسمها میشود. برای اینکه این مرور انجام گیرد مایل هستیم بر روی زیست شناسی تکاملی این پاتوژنهای قارچی تأکید شود و پتانسیل و احتمالاً مکانیسمهای تکاملی که به عملکردهای خاص ژنها و محصولات آنها منجر میشود مورد توجه قرار گیرد، بنابراین به جای تعریف کلمههای virulence factorو virulenceو بازتعریف آنها برای رسیدن به مفاهیم جدید پاتوژنز بیماریهای قارچی انسانی، استفاده از لغات جدیدی را توصیه میکنیم که منجر به درک صحیح زیست شناسی این پاتوژنهای قارچی بعنوان ارگانیسمهای ساپروفیتیک میشود.
به طور کلی این اصطلاحات باید بیانگر این موضوع باشند که توانایی اختصاصی برخی از قارچها در ایجاد بیماری در بسیاری از مواقع بستگی به وجود نقص در سیتم ایمنی میزبان دارد.
پیشنهاد میکنیم که قارچهای فرصت طلب بیماریزا برای انسان به جای (فاکتورهای ویرولانس کلاسیک) دارای (نشانها یا خواص ویرولانس) هستند. خواص ویرولانسی، اجزایی از این قارچها هستند که از طریق فشارهای انتخابی در طول دوره تکاملی بوجود آمدهاند تا به این قارچها اجازهی تطابق با محیطهای طبیعی و تکامل چرخهی زندگیشان را بدهند. به طور اتفاقی این نشانها یا خواص ویرولانسی به قارچهای خاصی این توانایی را داده است که در میزبانهایی که از نظر سیستم ایمنی دارای اختلال هستند، بتوانند تطابق حاصل کرده و زنده بمانند. اینچنین محیطهائی اغلب اوقات بیشباهت با آشیانههای اکولوژیکال طبیعی آنها نیست.
در ادامه بحث خود را بیشتر بر روی سه ارگانیسم قارچی مهم بیماریزا برای انسان یعنی آسپرژیلوس فومیگاتوس، کریپتوکوکوس نئوفرمنس و کاندیدا آلبیکنس متمرکز میکنیم و به طور خلاصه در مورد برخی از ویژگیهای قارچهای دوشکلی هم توضیحاتی ارائه خواهد شد. دراین بحثها ویرولانسهای اصلی این ارگانیسمها، (جدول 1) دلائل و مفاهیم مربوط به عفونتهای قارچی انسانی، مکانیسمهای تکاملی محتمل که بدان وسیله این خواص یا نشانهها ایجاد شدهاند و درمانهای احتمالی جدید که ممکن است با استفاده از این دانش جدید توسعه یافته باشد توضیح داده خواهد شد.
جدول 1: خواص عمومی مربوط به سه عامل پاتوژن قارچی شایع
| پاتوژن قارچی | آشیانه اکولوژیک | مرفولوژی | نشانهای ویرولانسی | بیماری |
| آسپرجیلوس فومیگاتوس | خاک، کود، مواد آلی در حال فساد، مخروبه | کنیدی، هایفی رشتهای | رشد در دمای بالا، مقاومت به استرس اکسیداتیو، رشد سریع، متابولیتهای ثانویه | آسپرجیلوز مهاجم، آسپرجیلوز برونکوپولمونری آلرژیک |
| کریپتوکوکوس نئوفرمنس | خاک، درختان، فضولات پرندگان | مخمر | رشد در دمای بالا، کپسول پلیساکاریدی، تولید ملانین | مننژیت کریپتوکوکی، بیماری ریوی |
| کاندیدا آلبیکنس | کومنسال انسانی | مخمر | رشد در دمای بالا، جذب و اتصال، تولید پروتئاز، تشکیل بیوفیلم، دیمورفیسم | عفونت سیستمیک، کاندیدیازیس دهانی، کاندیدیازیس واژینال |
آسپرژیلوس فومیگاتوس – کپک مخاطره آمیز
پیشرفتهایی که در تکنولوژیهای پزشکی حاصل شده سبب افزایش توانائی ما در درمان بیماریهای تهدید کننده حیات از جمله سرطان، لوسمی و یا نقص عضو شده است. هماکنون سالانه در حدود 15000 پیوند آلوژن و 25000 پیوند اتولوگ سلولهای بنیادی در سراسر دنیا انجام میپذیرد، به علاوه از سال 1988 تا 2002 در ایالات متحده آمریکا 113،682 پیوند عضو جامد صورت پذیرفته است که بیانگر افزایش 20% این امر در بازه زمانی 5 ساله میباشد. متأسفانه افرادی که تحت این روشهای نجاتبخش زندگی قرار میگیرند به سبب شرایط سرکوب ایمنی که لازم است تحمل کنند از ریسک بالای ابتلاء به عفونت با آسپرژیلوس فومیگاتوس و سایر گونههای آسپرژیلوسی برخوردار هستند. به طور کلی از 1980 به بعد میزان مرگ و میر ناشی از عفونت آسپرژیلوس فومیگاتوس 357% افزایش یافته است. از آنجائی که تکنولوژیهای پزشکی به پیشرفت خود ادامه میدهند و نیز به علت جمعیت رو به افزایش افراد دارای اختلال در سیستم ایمنی، بروز عفونتهای مهلک مربوط به آسپرژیلوس فومیگاتوس هم بسرعت (مثل موشک) رو به افزایش خواهد بود. اگرچه مطالعات اخیر در جمعیت در معرض خطر ابتلاء که در آن از آزولهای وسیعالطیف برای پیشگیری از عفونت قارچی استفاده شده، موفقیت آمیز بوده است، اما باید بطور کامل از عفونتهای آسپرژیلوسی در جمعیتهای ریسک پذیر جلوگیری بعمل آید.
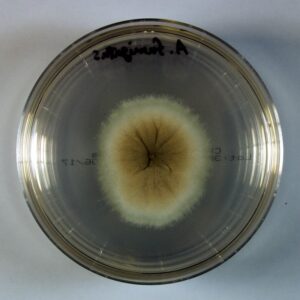
کلنی آسپرجیلوس فومیگاتوس

کلنیهای آسپرجیلوس فومیگاتوس
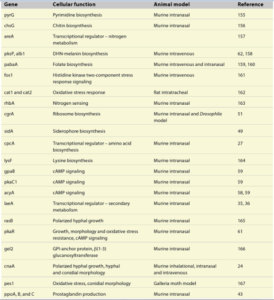
در رابطه با نیاز فوری برای درک مکانیسمهای بیماریزایی این قارچ رشتهای مهم، مطالعات ملکولی برای فهمیدن مکانیسمهای بیماریزایی آسپرژیلوس فومیگاتوس نیز افزایش یافته است. دستیابی کامل به توالی کل ژنوم استرین کلینیکی آسپرژیلوس فومیگاتوس و سایر گونههای مرتبط آسپرژیلوسی موجب تسریع در تحقیقات مربوط به پاتوژنز آسپرژیلوس فومیگاتوس شده است. در حال حاضر تحقیق بر روی مکانیزمهای مولکولی خواص بیماریزائی آسپرژیلوس فومیگاتوس در حال انجام است. تا این تاریخ هیچ فاکتور ویرولانسی در آسپرجیلوس فومیگاتوس شناسائی نشده است، اگرچه موتاسیون در حدود 20 ژن که همگی آنها در بیولوژی پایهی قارچ نقش دارند منجر به کاهش بیماریزایی این قارچ در مدل موشی آسپرژیلوزیس شده است. (جدول 2)
روی هم رفته این نتایج به ما این امکان را میدهد که نشانهای ویرولانسی منحصر به فردی که آسپرژیلوس فومیگاتوس را به یک قارچ کشنده فرصت طلب تبدیل کرده است را درک کنیم. این خواص یا نشانههای ویرولانسی تصور میشود که شامل ترموتولرنت بودن، سرعت رشد، مورفولوژی کنیدی، تولید متابولیتهای ثانویه، تولید آنزیمهای تجزیه کننده و مقاومت نسبت به استرسهای اکسیداتیو باشند. اگرچه همانطور که در جدول (2) هم نشان داده شده است اکثریت قریب به اتفاق ژنهایی که به عنوان ژنهای دخیل در پاتوژنز آسپرژیلوس فومیگاتوس معرفی شدهاند ژنهایی هستند که به طور مستقیم مربوط به متابولیسم اولیه ارگانیسم بوده و در نتیجه در رشد و نمو قارچ دخیل هستند، بنابراین تحقیقات بر روی بیماریزایی آسپرژیلوس فومیگاتوس تا حال حاضر قویاً مطرح میکند که آسپرژیلوس فومیگاتوس دارای فاکتورهای ویرولانس نیست بلکه دارای یک سری نشانهای ویرولانسی منحصر به فردی است که اجازه زنده ماندن در بدن میزبانان دچار نقص ایمنی را برایش فراهم میکند.
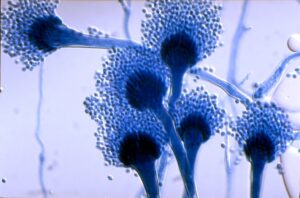
دستگاه زایشی آسپرجیلوس فومیگاتوس (منبع: اینترنت، دکتر میرهندی)
جدول 2: ژنهائی که در بیماریزايی آسپرجیلوس فومیگاتوس نقش دارند
پیشرفتهای جدید در درک مکانیسمهای بیماریزایی قارچهای فرصت طلب (2)
پیشرفتهای جدید در درک مکانیسمهای بیماریزایی قارچهای فرصت طلب (4)
پیشرفتهای جدید در درک مکانیسمهای بیماریزایی قارچهای فرصت طلب (3)
برای دانلود پی دی اف بر روی لینک زیر کلیک کنید
ورود / ثبت نام