متابولیسم کربن و انرژی باکتریهای بیهوازی
زکیه نظری، کارشناس ارشد میکروبیولوژی
Zakiye.nazari1987@gmail.com
متابولیسم انرژیزا در باکتریها
در باکتریهای مختلف برای تبدیل انرژی شیمیای یا انرژی نوری به نیرویی که بتواند از نظر بیولوژیک مورد استفاده قرار گیرد سه مسير تنفس، تخمیر و فتوسنتز موجود است. در تنفس مولکول اکسیژن آخرین پذیرنده الکترون است. در تخمیر مولکول غذایی معمولاً دو قسمت میشود که بعداً یکی بوسیله دیگری اکسیده میگردد. در فتوسنتز، انرژی نوری به انرژی شیمیایی تبدیل میشود. در تمام این مسيرهاي انرژیزا در تمام پدیدهها، آدنوزین تری فسفات (ATP) تشکیل میشود.
دسته بندی باکتریها از نظر نیاز به اکسیژن
باکتریهای هوازی اجباری: برای رشد احتیاج به اکسیژن آزاد دارند و زنجیره تنفسی انتقال الکترون آنها کامل است. باکتریهای بیهوازی اجباری تنها در شرایطی رشد میکنند که درجه احیا بسیار زیاد باشد و اکسیژن از محیط خارج شود، چون اکسیژن برای آنها سمی است و موجب مرگشان میشود.
باکتریهای بیهوازی مقاوم در برابر هوا: وجود اکسیژن هوا را تحمل میکنند و اگر در معرض آن قرار گیرند کشته نمیشوند.
باکتریهای بیهوازی اختیاری: در حضور اکسیژن و یا در غیاب آن رشد میکنند و بسته به شرایط، زندگی هوازی یا بیهوازی دارند.
باکتریهای میکروآیروفیلی: بهترین وضعیت رشد آنها در فشار کم اکسیژن است و اگر فشار اکسیژن زیاد باشد قادر به رشد نیستند. در باکتریهای بیهوازی اختیاری و بیهوازی مقاوم در برابر هوا، متابولیسم انرژیزا بطور مطلق از راه تخمیر انجام میشود. وقتی اجرام در حضور هوا کشت داده شوند تعدادی از واکنشهای آنزیمی جریان مییابند که به تولید هیدروژن پراکسید و رادیکال سوپراکسید میانجامد که هر دو برای زندگی یاختهای خطرناکند. در باکتریهای هوازی، بیهوازی اختیاری و بیهوازی مقاوم در برابر هوا آنزیم سوپر اکسید دیسموتاز از تجمع یون سوپر اکسید جلوگیری میکند ولی در باکتریهای بیهوازی اجباری چنین آنزیمی وجود ندارد. هیدروژن پراکسید که بوسیله واکنش دیسموتاز تولید میشود سریعاً بوسیله آنزیم کاتالاز موجود در باکتریهای هوازی و بیهوازی اختیاری تخریب میشود.
تخمیر و تنفس در باکتریها
باکتریهایی که از انرژی شیمیایی استفاده میکنند دو مکانیسم اساسی تأمین انرژی یعنی تخمیر و تنفس را به کار میگیرند. در پدیده تخمیری، الکترونها از دهنده الکترون یعنی یک ماده واسطهای ناشی از شکستن مولکول ماده اولیه به پذیرنده الکترون متصل میشوند که ماده واسطهای دیگری در پدیده تخمیر است. تخمیر بوسیله باکتریهای هوازی اجباری و اختیاری انجام میشود و برای انجام آن وجود اکسیژن لازم نیست. تنفس پدیدهای است که در آن معمولاً مولکول اکسیژن به عنوان پذیرنده الکترون بکار میرود. وقــتی اکسیژن پذیرنده نهایی باشد، پدیده را تنفس هوازی مینامند ولی در تنفس بیهوازی یک ترکیب غیر آلی از قبیل نیترات، سولفات و يا کربنات دریافت کننده الکترون است.
تخمیر و تنفس در باکتریها
در طی فرمانتاسیون، محصولات واسطهای که در اثر کاتابولیسم یک ماده آلی مثل گلوکز بوجود میآیند، به عنوان یک ماده گیرنده نهایی الکترون عمل میکنند و در نتیجه یک محصول پایدار از تخمیر بوجود میآید.
تخمیر در مقایسه با تنفس
تخمیر در مقایسه با تنفس انرژی کمتری از مولکول ماده اولیه استخراج میکند. وقتی اجرام گلوکز را تخمیر میکنند، تنها قسمتی جزئی از توان انرژی که در گلوکز نهفته است آزاد میشود و اکثر انرژی این ماده در فراوردههای این واکنش از قبیل لاکتات محبوس میماند، ولی وقتی اجرام گلوکز را اکسیده میکنند و H2O و CO2ایجاد میشود، تمام انرژی قابل دسترس مولکول گلوکز آزاد میگردد. تخمیر معادل 47 کیلوکالری بر مول و اکسیداسیون 686 کیلوکالری بر مول انرژی تولید میکند، پس باکتریهای بیهوازی مطلق که از راه تخمیر کسب انرژی میکنند احتیاج به غذای بیشتری دارند.
متابولیسم کربن و انرژی در باکتریهای بیهوازی
گلوکز در متابولیسم انرژیزا از اهمیت خاصی برخوردار است. در اکثر اشکال زندگی گذرگاه متابولیسمی خاصی برای آن وجود دارد. یاخته ابتدا یک قند یا ترکیب وابسته به آن را به مواد واسطهای مشترک گذرگاه تخمیری گلوکز تبدیل میکند و سپس مورد استفاده قرار میدهد.
گذرگاه گلیکولیتیک
در اکثر یاختهها مهمترین راه کاتابولیسم گلوکز همانا گلیکولیز است که در جریان آن مولکول گلوکز بدون دخالت اکسیژن به دو مولکول اسید لاکتیک تجزیه میشود. اساس گلیکولیز در واکنشهای یازده آنزیمی امبدن- میرهف پارناز(EMP) است. راه گلیکولیز به نام دو دانشمندی که در مشخص کردن این راه بسیار کوشیدهاند به نام راه امبدن- میرهف نیز معروف است.
گلیکولیز اصولاً متشکل از دو مرحله عمده است:
مرحله اول: گلوکز بسته به جرم بوسیله ATP یا PEP فسفریله شده و شکاف میخورد تا گلیسرآلدهید 3- فسفات تشکیل شود.
مرحله دوم: در این مرحله انرژيي که در اصل در مولکول گلوکز نهفته است به شکل ATP نگهداری میشود.
مراحل دهگانه گلیکولیز
مرحله اول
ورود D-گلوکز در راه گلیکولیز مستلزم فسفریلدار شدن آن به گلوکز 6- فسفات است که بوسیله آنزیم هگزوکیناز کاتالیز میشود. این آنزیم، آنزیم اختصاصی است که گلوکز را در کربن ششم فسفریلدار میکند. بدین منظور مولکولATP آبکافت شده و مقداری انرژی برابر 7/3کیلوکالری بر مول آزاد میسازد. از این مقدار انرژی 3/3 کیلوکالری آن به مصرف تشکیل پیوند فسفو گلوکز میرسد و بقیه ذخیره میشود در نتیجه مقدار انرژی آزاد شده 4 کیلوکالری بر مول است.
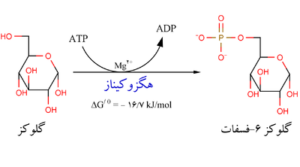
مرحله دوم
تبدیل گلوکز 6- فسفات به فروکتوز 6- فسفات است. این واکنش یک واکنش ایزومری شدن است که بوسیله آنزیم فسفو هگزوز ایزومراز کاتالیز میشود.
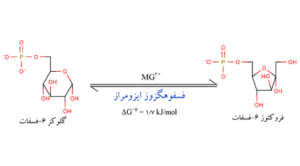
مرحله سوم
فسفریلدار شدن مجدد فروکتوز 6- فسفات بوسیله آنزیم 6- فسفوفروکتوکیناز است. در این حالت، آنزیم گروه فسفات حاصل از آبکافت،ATP را به مولکول فروکتوز 6- فسفات انتقال میدهد و در نتیجه فروکتوز 6،1- دیفسفات حاصل میشود.
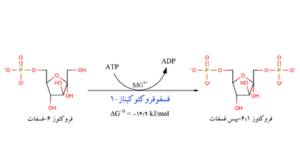
مرحله چهارم
مرحلهای است که طی آن گلیسرآلدهید 3- فسفات ساخته میشود. مولکول فروکتوز 6،1 دیفسفات بوسیله آنزیم آلدولاز به دو ترکیب سه کربني دیهیدروکسی استون فسفات و گلیسرآلدهید 3- فسفات تخریب میشود. دیهیدروکسی استون فسفات به نوبه خود میتواند تحت تأثیر آنزیم تریوز فسفات ایزومر به گلیسرآلدهید 3- فسفات تبدیل گردد.
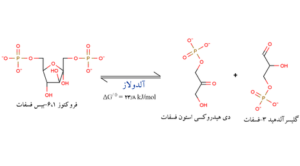
مرحله پنجم
به مرحله اکسایش گلیسرآلدهید 3- فسفات معروف است. در این مرحله گلیسرآلدهید 3- فسفات تحت تأثیر آنزیم گلیسرآلدهید 3- فسفات دهیدروژناز به 3،1- دیفسفوگلیسریک اسید تبدیل میگردد.
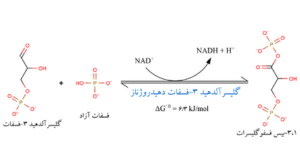
مرحله پنجم
این واکنش نیازمند کوآنزیم نیکوتین آمید آدنین دینوکلئوتید و فسفات کانی است. واکنش در دو مرحله انجام میگیرد که یکی انرژیزا و دیگری انرژیخواه است. ابتدا گلیسرآلدهید 3- فســفات تحت تأثیر NAD+ اکسید شده و 1- فسفوگلیسریک اسید میدهد که واکنش انرژیزا است، سپس این ترکیب بوسیله فسفات کانی فسفریلدار شده و 3،1- دیفسفوگلیسریک اسید را میسازد که انرژیگیر است.
مرحله ششم
این مرحله یکی از مهمترین مراحل راه گلیکولیز است، زیرا نخستین مرحلهای است که طی آن یک مولکول پر انرژی از نوعATP سنتز میشود. برای تشکیل مولکولATP ، حداقل انرژی برابر 7/3 کیلوکالری بر مول لازم است. از برداشت گروه فسفات متصل به کربن شماره 1 ترکیب 7/3 کیلوکالری بر مول به مصرف تشکیلATP ميرسد و 4/5 کیلوکالری بر مول باقی میماند.
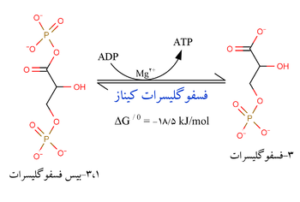
مرحله هفتم
واکنش سادهای است که بوسیله آنزیم فسفوگلیسرو موتاز کاتالیز میشود. در این حالت، فسفات متصل به کربن شماره 3 ترکیب 3- فسفوگلیسرات، به کربن شماره 2 منتقل شده و 2- فسفوگلیسرات میدهد.
مرحله هشتم
این واکنش یک واکنش آبگیری از 2- فسفوگلیسرات است که توسط آنزیم آنولاز کاتالیز میشود. طی این واکنش، 2- فسفوگلیسرات به فسفوانول پیرووات که ترکیبی پر انرژی است تبدیل میگردد.
مرحله نهم
این واکنش نیز یکی دیگر از واکنشهای مهم راه گلیکولیز است که طی آن دومین مولکول پر انرژی ATP سنتز میشود. آنزیم پیرووات کیناز واکنش را کاتالیز میکند. فسفات متصل به کربن شماره 2 فسفوانول پیرووات به مولکولADP منتقل وATP تشکيل میشود. فسفوانول پیرووات ترکیب پر انرژی است و این انرژی در پیوند فسفات متصل به کربن شماره 2 نهفته است. در اثر برداشت این فسفات، مقداری انرژی برابر 14/8 کیلوکالری بر مول تولید میشود که 7/3 کیلوکالری آن صرف ساخته شدنATP شده و بقیه ذخیره میگردد.
مرحله آخر
آخرین مرحله راه گلیکولیز واکنشی است که طی آن پیرووات احیا شده و لاکتات تولید میشود. آنزیم لاکتات دهیدروژناز و کوآنزیمNADH واکنش را کاتالیز میکنند.
نتیجه
راه گلیکولیز مکانیسم بیوشیمیایی است که از آن طریق انرژی شیمیایی گلوکز دوباره در سایر فرایندهای بیوشیمیایی مورد استفاده قرار میگیرد. از جمع بندی ترکیبات مصرف شده و مواد تولید شده معلوم میشود که تا مرحله تولید گلیسرآلدئید 3- فسفات دو مولکولATP مصرف میشود. از سوی دیگر در تبدیل دو مولکول گلیسرآلدئید 3- فسفات به پیرووات نیز چهار مولکولATP تولید میگردد. با کم کردن تعداد ATP مصرف شده از تعداد تولید شده، میزان کل انرژی حاصل از راه گلیکولیز دو مولکولATP خواهد بود.
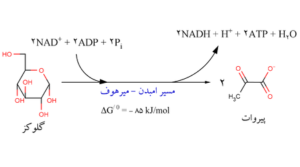
مسیر امبدن- میرهف

گذرگاه فسفوگلوکنات
(پنتوز فسفات، شانت هگزوز مونوفسفات)
مسیری است که هگزوزها و پنتوزها را به یکدیگر تبديل میکند. این مسیر که در بیشتر اندامها رخ میدهد به عنوان منبع تولید کاهنده NADPH و پنتوز برای فرایندهای بیوسنتزی عمل میکند. باکتریهایی که تخمیر هترولاکتیک دارند و همچنین بروسلا آبورتوس از این گذرگاه استفاده میکنند. وقتی گلوکز از راه فسفوگلوکنات تخمیر میشود، میزان ATP حاصله نصف ATPناشی از گذرگاه EMP است.
گذرگاه فسفو گلوکنات
NADPH آن در بیوسنتزهای احیایی استفاده میشود، در صورتی که ریبوز 5- فسفات در سنتز RNA و DNA و کوآنزیمهای نوکلئوتیدی مورد استفاده قرار میگیرد.
مسیر پنتوز فسفات دو فاز دارد:
NADPH فاز اول اکسیداتیو و فاز دوم غیر اکسیداتیو است. اولین واکنش این مسیر دهیدروژناسیون گلوکز 6- فسفات توسط گلوکز 6- فسفات دهیدروژناز و تولید 6- فسفوگلوکنولاکتون میباشد و به عنوان گیرنده الکترون عمل میکند. این لاکتوز توسط یک لاکتوناز اختصاصی به اسید آزاد 6- فسفوگلوکنات هیدرولیز شده است.
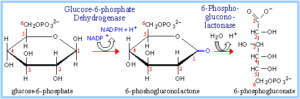
6- فسفو گلوکنات خود طی دهیدروژناسیون و کربوکسیلاسیون توسط 6- فسفوگلوکنات دهیدروژناز به کتوپنتوز D-ریبولوز 5 فسفات تبدیل میگردد. طی این واکنش مولکول دومNADPH تولید میشود.

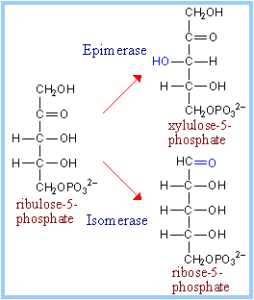
با فعالیت آنزیم فسفو پنتوز ایزومراز، ریبولوز 5- فسفات به ایزومر آلدولاز یعنی D-ریبوز 5- فسفات تبدیل میگردد. در بعضی بافتها این مسیر در این نقطه به پایان میرسد. در بافتهایی که اساساً تنها نیاز به NADPH دارند، ریبوز 5- فسفات طی یک سری از واکنشها مجدداً به گلوکز 6- فسفات تبدیل میگردد.
در قسمت غیر اکسیداتیو مسیر پنتوز فسفات، ترانس کتولاز انتقال یک قطعه دو کربنه گزیلولوز 5- فسفات را به ریبوز 5- فسفات کاتالیز نموده و تولید سودوهپتولوز 7- فسفات مینماید.
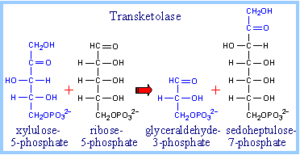
قطعه سه کربنه باقیمانده گزیلولوز، گلیسرآلدئید 3- فسفات میباشد. ترانس آلدولاز با برداشت یک قطعه 3 کربنی از سودوهپتولوز و ترکیب آن با گلیسرآلدئید 3- فسفات، تولید فروکتوز 6- فسفات مینماید. قطعه 4 کربنه باقیمانده از سودو هپتولوز، اریتروز 4- فسفات خواهد بود.
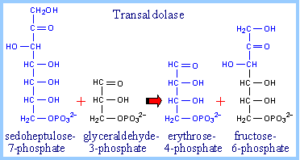
با ترانس کتولاز از اریتروز 4- فسفات و گزیلولوز 5- فسفات تولید فروکتوز 6- فسفات و گلیسرآلدئید 3- فسفات میگردد. دو مولکول گلیسرآلدئید 3- فسفات حاصل از این واکنشها میتوانند به یک مولکول فروکتوز 1،6 بیسفسفات تبدیل شوند.آنزیم ترانس کتولاز و ترانس آلدولاز که دو آنزیم مهم این مسیر هستند، ارتباطی برگشت پذیر بین مسیر پنتوز فسفات و گلیکولیز برقرار میکنند.
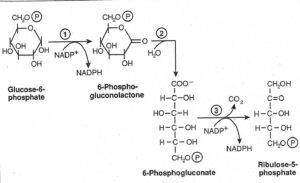
گذرگاه انتنر دودراف
این مسیر اغلب در سودوموناس، ریزوبیوم، ازتوباکتر، آگروباکتر و مقدار کمی از دیگر باکتریهای گرم منفی و همچنین در تعدادی از گرم مثبتها مثل انتروکوکوس فکالیس دیده میشود. این گذرگاه از ناحیه 6- فسفو گلوکنات از گذرگاه فسفو گلوکنات منشعب میشود. در این ردیف واکنش، 6 فسفو گلوکنات بدون آب میشود و بعداً به یک مولکول گلیسرآلدئید 3- فسفات و یک مولکول پیرووات تبدیل میگردد. از پیرووات، الکل اتیلیک (اتانول) و CO2 بوسیله واکنشهایی تولید میگردد که در مورد تخمیر الکلی مخمرها صادق است. در این گذرگاه نیز همانند گذرگاه فسفوگلوکنات به ازای تخمیر یک مولکول گلوکز، تنها یک مولکول ATP ايجاد میشود، همچنین بازده این مسیر یک مولکولNADH و یک مولکولNADPH هم هست.
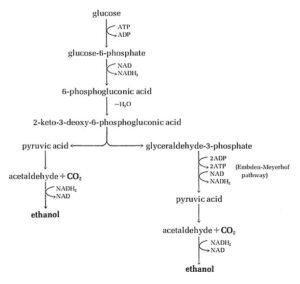
این فرایند تنها در پروکاریوتها انجام میشود. در برخی اجرام، آخرین فرآورده، الکل و یا اسید لاکتیک است. در برخی دیگر از باکتریها، اسید پیروویک بیشتر متابولیزه میشود و به فرآوردههایی از قبیل بوتیریک اسید، بوتیل الکل، استن یا پروپیونیک اسید مبدل میشود.
تخمیر الکلی
قدیمیترین نوع شناخته شده تخمیر الکلی، تولید اتانول از گلوکز است. در مخمرها که تخمیر الکلی تقریباً بطور خالص انجام میشود، پیرووات دکربوکسیلاز که آنزیم کلیدی تخمیر الکلی است، اسید پیروویک را دکربوکسیله کرده و منشأ تولید الکل میگردد. در این جریان استالدئید آزاد تشکیل میشود که بعداً بوسیله الکل دهیدروژناز، احیا شده و به اتانول تبدیل میشود. در این واکنشNADH به عنوان دهنده الکترون است. اتانول و CO2 تولید شده در تخمیر باعث ورآمدن خمیر نان و مطلوب شدن کیفیت نان میشود.
- C6H12O6 → 2C2H5OH + CO2
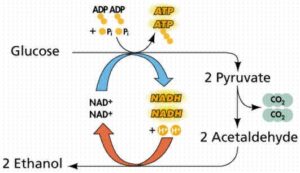
تخمیر اسید لاکتیک
تخمیر اسید لاکتیک، احیاء پیرووات به لاکتات است. در باکتریها (باکتریهای اسید لاکتیک و باسیلوس)، جلبک (کلرلا)، بعضی کپکهای آبزی و پروتوزوآ و حتی در عضله حیوانات به شکل معمول وجود دارد.
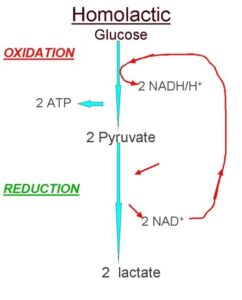
تخمیر همولاکتیک
تمام اعضای گروه استرپتوکوکوس و بسیاری از انواع لاکتوباسیلوس گلوکز را عمدتاً به اسید لاکتیک تخمیر میکنند.
سایر فرآوردههای تخمیری دیگر بصورت ناچیزی تولید میشود. از آن نظر که در این نوع پدیده تنها اسید لاکتیک بدست میآید، نوع تخمیر آن را همولاکتیک و باکتریهای تخمیر کننده را هموفرمانتر مینامند.
تخمیر هترولاکتیک
در برخی از باکتریهای اسید لاکتیک از قبیل لکونوستوک و بعضی از لاکتوباسیلوسها، پدیده تخمیری حالت مخلوط دارد.
تنها تقریباً نیمی از گلوکز را به اسید لاکتیک تبدیل میکنند و بقیه به co2 ،الکل، اسید فورمیک یا اسید استیک مبدل میشوند، به همین علت است که اینگونه تخمیر را هترولاکتیک مینامند. از گذرگاه پنتوز فسفات بیشتر استفاده شده است. در تخمیر هترو لاکتیک، میزان انرژی حاصل از یک مولکول گلوکز، یک سوم انرژی ناشی از تخمیر همولاکتیک است.
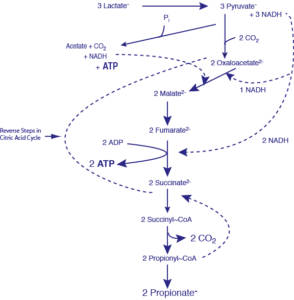
تخمیر اسید پروپیونیک
یکی از فرآوردههای انتهایی مشخص و مهم تخمیر برخی از باکتریهای بیهوازی، پروپیونات است. وجه مشخصه اجرام گونه پروپیونیباکتریوم و باکتری بیهوازی میلهای شکل، گرم مثبت وابسته به لاکتوباسیلهاست. بو و طعم خاص پنیر سویسی که یکی از منابع این نوع باکتریهاست، بواسطه وجود اسید پروپیونیک است. کوآنزیم این تخمیر بیوتین است. انرژی موجود در پیوند پروپیونیل کوآنزیم A فقط از طریق واکنش پروپیونیل کوآنزیم A با اسید سوکسینیک برای تشکیل سوکسینیل- کوآنزیم A و اسید پروپیونیک آزاد تشکیل میشود. بعلاوه CO2 آزاد شده از دکربوکسیلاسیون متیل مالونیل کوآنزیم A بصورت پیوند با آنزیم دارای بیوتین باقی میماند و بطور مستقیم CO2 را به اسید پیروویک منتقل میکند و اگزالو استیک تشکیل میشود.
تخمیر فرمیک اسید
خیلی از باکتریها مخصوصاً تعدادی از خانواده انتروباکتریاسه میتوانند پیرووات را به اسید فرمیک و دیگر محصولات متابولیزه کنند. گاهی به این پروسه تخمیر اسید فرمیک گویند. فرمیک اسید ممکن است بوسیله فرمیک هیدروژنیلاز به H2 و CO2 تبدیل شود. دو نوع تخمیر اسید فرمیک داریم:
- تخمیر اسیدهای مخلوط
- تخمیر بوتاندیول
تخمیر اسیدهای مخلوط
باکتریهای گونههای اشریشیا، سالمونلا و شیگلا قندها را از راه پیرووات به اسیدهای لاکتیک، سوکسینیک، استیک و فرمیک تبدیل میکنند، علاوه بر این Co2 و H2 و اتانول نیز تولید میشود. تمام انتروباکتریاسهها فرمات میسازند که آن را یا انباشته کرده و یا در شرایط اسیدی بوسیله فرمیک هیدروژنیلاز به هیدروژن و کربن اکسید میکند. برای تشکیل اسید فرمیک، کوآنزیم A، پیرووات را میشکافد تا استیل کوآنزیم A و فرمات تولید شود. استیل کوآنزیم A سریعاً به استیل فسفات مبدل میگردد. وجه مشخصه تخمیر اشریشیا کلی و اکثر سالمونلاها تولید CO2 و H2 است، در حالی که شیگلا و سالمونلا تیفی CO2 و H2 تولید نمیکنند و معادل این مواد، اسید فرمیک انباشته میشود. مسئول واکنش متیل-رد فقط برای تخمیر اسیدهای مخلوط مثبت است، چون PH به 5/4 میرسد و رنگ شاخص از زرد به قرمز تبدیل میشود.

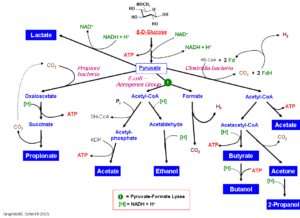
تخمیر بوتاندیول
مشخصه انتروباکتر، سراشیا، اروینیا، باسیلوس و کلبسیلا است. دو مولکول پیرووات به استوئین تبدیل میشود که سپس با NADH تبدیل به 2 و 3 بوتاندیول میشود. این واکنش احیا در مجاورت هوا به آهستگی قابل برگشت است و وقتی محیط شدیداً قلیایی شود واکنش Voges-Proskauer را تشکیل میدهد که یک آزمایش جستجوی استوئین است.

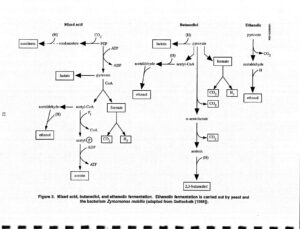
تخمیر بوتیریک اسید
در بسیاری از باکتریهای گونههای کلستریدیوم، بوتیریک اسید، استیک اسید، CO2 و H2 مهمترین فرآوردههای مشخص تخمیر کربوهیدراتها هستند. واکنش کلیدی، تشکیل استواستیل- کوآنزیم A است که از تغلیظ دو مولکول استیل کوآنزیم A حاصل میشود که خود مشتق از استات یا پیرووات میباشد. این ترکیب چهار کربنی بعداً احیاء میشود و به اسید بوتیریک تبدیل میگردد و ATP تشکیل میشود. در برخی از اجرام اولین فرآوردههای اسیدی تخمیر، احیا میشوند و به انباشتگی فرآوردههای انتهایی خنثی منجر میگردند که عبارتند از: بوتانول، استون، ایزوپروپانول و اتانول.
کلستریدیوم استوبوتیلیکوم یک باکتری انتخابی برای تخمیر استون- اتانول- بوتانول است. باکتریهای فوق در جریان تخمیر سه دسته فرآورده تولید میکنند:
- استون- بوتانول- اتانول
- اسیدهای آلی استیک، بوتیریک و لاکتیک
- CO2 و H2
انتقال الکترون در باکتریهای بیهوازی
در باکتریهای هوازی اجباری اکسیژن تنها پذیرنده انتهایی الکترون است و سیتوکرومهای نوع a ، d و … به عنوان اکسیدازهای انتهایی فعالیت میکنند. در اجرام بیهوازی اختیاری تعداد متنوعی از سایر پذیرندههای الکترون دخالت دارند. وقتی این باکتریها در شرایط بیهوازی رشد میکنند، دستگاه انتقال الکترون آنها ممکن است با پذیرندههای غیر از اکسیژن همگام شود که از آن جمله نیترات، سولفات و فومارات را میتوان ذکر کرد. تولید ATP در جوار بسیاری از واکنشهای پذیرنده الکترون و مصرف هیدروژن چه در بیهوازی اختیاری و چه در بیهوازی اجباری خودنمایی میکند. تنفس ارگانیسمهای بیهوازی را به سه گروه فرعی تقسیم میکنند: احیاء کننده نیتراتها، احیاء کننده سولفاتها و باکتریهای متان.
احیای نیترات به نیتریت
مشخصترین دستگاه الکترونی بیهوازی، تنفس نیترات است كه نوعی تنفس بیهوازی است و در آن نیترات آخرین پذیرنده الکترون است. احیای نیترات به نیتریت بوسیله یک دستگاه انتقال الکترون انجام میشود. در اشریشیا کولی که در شرایط بیهوازی در مجاورت نیترات کشت داده شده، فورمات مؤثرترین دهنده الکترون است. از جمله سایر مواد اولیهای که ممکن است به عنوان دهنده الکترون عمل کنند، لاکتات، سوکسینات و NADH قابل ذکرند.
نیتریت که فرآورده ناشی از تنفس نیترات است، فوقالعاده سمی است و در مجاورت آن رشد اکثر اجرام محدود میشود، با وجود این در معدودی از اجرام از قبیل باسیلوس و سودوموناس، نیترات ممکن است احیا شود که از مرحله نیتریت بگذرد و نیتروژن مولکولی تولید گردد. چنین واکنشي که نیتریتزدایی (دنیتریفیکاسیون) نامیده میشود، بوسیله ردیفی از واکنشهای تنفس بیهوازی انجام میگیرد. باکتریهایی که قادرند نیترات را به جای اکسیژن استفاده کنند عبارتند از: باسیلوس، سودوموناس، اشریشیا، تیوباسیلوس دینیتریفیکان، انتروباکتر، میکروکوکوس و ریزوبیوم.
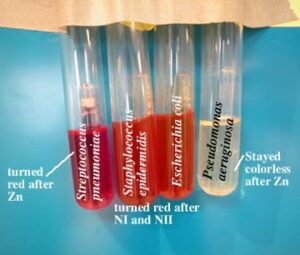
احیاء کنندههای سولفات
دی سولفوویبریو و دیسولفوتوماکولم که اندوسپور تولید میکند، ارگانیسمهای بیهوازی اجباری هستند که سولفات را به عنوان گیرنده نهایی الکترون استفاده میکنند و به کمک آنها سولفات احیاء شده تبدیل به سولفیت میشود. چون این ارگانیسمها به یک منبع آلی کربن احتیاج دارند، از این رو هتروتروف هستند.
باکتریهای متانوژنیک
متان احیاء شدهترین ترکیبات آلی است و در آخرین مرحله یکی از زنجیرههای غذایی بیهوازی تشکیل میشود. متان عمدتاً از استات، CO2 و H2 بوسیله باکتریهای بیهوازی متانوژن تولید میشود. این باکتریها در محیط بیهوازی محلهايي زندگی میکنند که در آن مواد آلی در حال تجزیهاند. متانوژنها برای رشد به شرایط بیهوازی بسیار شدیدی نیازمندند. متانزایی آنها تنها وقتی است که ردوکس پتانسیل کمتر از 230 میلیولت باشد.
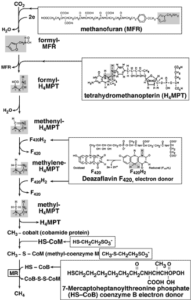
این باکتریها برای تنفس بیهوازی از کربن دیاکسید به عنوان پذیرنده اصلی الکترون استفاده میکنند. این باکتریها از دو دسته باکتریهای مزوفیل و گرمادوست هستند و به ترتیب در دمای 37 تا 40 درجه سانتیگراد و 50 تا 52 درجه سانتیگراد میتوانند زندگی کنند. مواد واسطهای در احیای CO2 به متان، فورمات، فرمالدئید و متانول است. یکی از حاملین کوآنزیم M است که ماهیت شیمیایی آن 2- مرکاپتواتان سولفونیک اسید است.
بیوگاز
مخلوطی ازسه ترکیب عمده به نامهای متان، دیاکسید کربن و سولفید هیدروژن است که حاصل تجزیه بیهوازی و تخمیر باکتریهای متانوژن است. جزء مسئول اشتعال بیوگاز، متان است. متان گازی بیرنگ و بیبو است که اگر یک فوت مکعب آن بسوزد، 252 کیلو کالری انرژی حرارتی تولید میکند. از مزیتهای مهم متان به دیگر سوختها این است که هنگام سوختن گاز سمی و خطرناک منوکسید کربن تولید نمیکند، بنابراین از آن میتوان به عنوان سوخت ایمن و سالم در محیط خانه استفاده کرد. شرایط مطلوب تولید بیوگاز دمای 70 درجه سانتیگراد است که باکتریها بیشترین فعالیت آنزیمی را برای تجزیه مواد آلی و تولید بیوگاز دارند و همچنين قلیایی بودن (8-7pH ) محیط واکنش است.
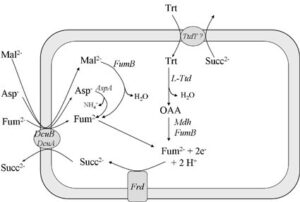
احیای فومارات
تعدادی از باکتریهای بیهوازی مطلق از فومارات به عنوان یک پذیرنده الکترون استفاده میکنند. فومارات به آسانی از منابع مختلف کربن مثل مالات، آسپارتات و پیرووات تولید میشود. در اشریشیا کلی که در شرایط بیهوازی در محیط گلیسرول همراه با فومارات به عنوان پذیرنده نهایی الکترون بوجود آمده، وجود سیتوکروم به اثبات نرسیده است كه این امر نشانهای است از تخریب سیتوکروم در جریان تنفس بیهوازی نوع فومارات.
باکتریهای متیلوتروف
انرژی مورد نیاز خود را از اکسیداسون گروههای متیلی به دست میآورند. برخی از این باکتریها اجباریاند و تنها در ترکیباتی بكار میروند که اتصال کربن-کربن ندارند (متان و متانول). اکسیداسیون متان به کربن دیاکسید از طریق مراحل مختلف الکترون- اکسیداسیون انجام میگیرد. در این واکنشها فرمالدئید وضعیت کلیدی دارد.
منابع:
- Microbioligy; lansing M.Prescott
- باکتری شناسی عمومی، دکتر حسن تاجبخش، انتشارات دانشگاه تهران
- اصول بیوشیمی لنینجر، نلسون کاکس، ترجمه دکتر رضا محمدی
- میکروبیولوژی عمومی، انتشارات دانشگاه گیلان
- میکروب شناسی پزشکی جاوتز
https://medlabnews.ir/%d9%85%d8%ad%db%8c%d8%b7%e2%80%8c%d9%87%d8%a7%db%8c-%da%a9%d8%b4%d8%aa/
برای دانلود پی دی اف بر روی لینک زیر کلیک کنید
ورود / ثبت نام