نقش ترانسپوزون ها در پروکاریوتها (قسمت اول)
وهاب پیرانفر (کارشناس ارشد)، محمد عرفانی (کارشناس ارشد)، دکتر رضا میرنژاد (دانشیار دانشگاه)
مقدمهای بر ترانسپوزون ها[1]
از 1930 میلادی، مطالعات ژنتیکی بهصورت مستقل توسط باربارا مککلینتوک[2] و مارکوس روداس[3] روی گیاه ذرت انجام گرفت. نتایج این مطالعات آشفتگی زیادی در مورد ژنهای ساکن در جایگاههای ثابت درکروموزمهای اصلی به همراه داشت. گزارشهای این دو دانشمند نشان میداد که عناصر ژنتیکی بر روی کروموزوم اصلی وجود دارند که میتوانند از یک موقعیت به موقعیت دیگری نقلمکان کنند. هرچند این یافتهها تا سالها مورد شک و تردید قرار داشت، اما در حال حاضر روشن است که چنین عناصری همواره در طبیعت وجود داشتهاند. امروزه میدانیم که کروموزومهای پروکاريوتي، ویروسها و ژنوم يوکاريوتي حاوي قطعاتي از DNA هستند که میتوانند حرکت کرده و به مکانهای متفاوتي از همان کروموزوم و یا کروموزوم دیگری مهاجرت کنند. اين حرکت، جابجایی (ترانسپوزیشن[4]) خوانده میشود و نقش مهمي در تولید ترکيبات جديد ژني بازي میکند.
قطعات DNA که ژنهای موردنیاز براي جابجايي را حمل میکنند عناصر قابلانتقال يا ترانسپوزون هستند و برخي اوقات “ژنهای جهش دهنده” ناميده میشوند، اما نامهای بسیاری (که بعضی برای توصیف عمل آنهاست) بر این عناصر ژنتیکی نهادهاند، مانند عناصر کنترلکننده، cassettes، ژنهای پرشی[5]، ژنهای سیار[6]، ژنهای همراه، عناصر ژنتیکی همراه[7] و ترانسپوزون ها. ما اصطلاح “عناصر ژنتیکی قابل جابجایی“ را که بهطور رسمی درست و شامل تمام اعضای خانواده است را انتخاب میکنیم. اصطلاح جابجایی (ترانسپوزیشن) بهطور مدید در علم ژنتیک مورداستفاده قرار گرفته است و مشخصاً برای انتقال بخشی از کروموزوم از یک موقعیت به موقعیت دیگر و بازآرایی ساختار آن به کار میرود.
قطعات DNA که ژنهای موردنیاز براي جابجايي را حمل میکنند عناصر قابلانتقال يا ترانسپوزون هستند و برخي اوقات ژنهای جهشدهنده ناميده میشوند. الحاق این عناصر ژنتیکی نهتنها عملکرد ژن در منطقهای که در آن قرار میگیرد را از بين میبرد، بلکه میتواند عملکرد دوگانه داشته باشد يعني روي بيان ژن پاييندست اثر بگذارد؛ در حقيقت ترانسپوزون ها مانند موتاژنها عمل میکنند. در حال حاضر یک قطعه کوچک ژنی یا تعداد کمی از آنها که ارتباط خاصی در عملکرد با قابلیت جابجایی دارند در این خانواده قرار میگیرد.
جابجایی این قطعات میتواند روی همان کروموزوم و یا بر روی یک کروموزوم جدید باشد. قوانین و ارتباطاتی که این موضوع را سبب میشود در دست مطالعه است. این قطعات فقط از لحاظ ژنتیکی و فیزیکی شناسایی شدهاند. بررسی ایجاد ناهنجاری در فعالیت و ساختارهای ژن که در محل ورود این قطعات قرار دارد، از راههای شناسایی ژنتیکی آنهاست. متدهایی از قبیل تعیین توالی رشتههای الگو نیز مورد کاوش قرار گرفتهاند که از روشهای فیزیکی محسوب میشوند. نتایج نشان داده است که این عناصر جابجاشونده در اکثر موجودات زنده روی کره زمین وجود دارند.
امروزه این عناصر کلیدهای ژنتیکی ارزشمندی در پروکاریوتها و یوکاریوتها محسوب میشوند. با استفاده از این عناصر، نقشهبرداری ژنتیکی، ایجاد جهش، کلونسازی ژنها و حتی تولید موجود تراریخته زنده با سهولت بیشتری صورت میگیرد.

شکل 1) باربارا مک کلینتوک – دریافت جایزه نوبل در 1985 جهت توضیح رنگارنگی دانههای ذرت و کشف ترانسپوزونها
توالیهای الحاقشونده در باکتریها
توالیهای الحاقی (IS)[8] در حال حاضر به خوبی شناخته شدهاند؛ آنها بخشی از DNA باکتریایی هستند که میتوانند از موقعیتی به موقعیت دیگری بر روی یک کروموزم و یا یک محل جدید انتقال یابند. الحاق آنها نهتنها عملکرد ژن در منطقهای که در آن قرار میگیرند را از بين میبرد، بلکه میتواند عملکرد دوگانه داشته باشد؛ يعني روي بيان ژن پاييندست اثر بگذارد. در حقيقت ترانسپوزون ها مانند موتاژنها عمل میکنند. اگر آنها به وسط یک ژن منتقل شوند، کد قطعه ژنتیکی را بیمعنی و حتی از بیان آن جلوگیری میکنند. آنها همچنین با در نظر گرفتن طولشان میتوانند حاوی کدهای پایان ترجمه و رونویسی باشند و از بیان سایر ژنها در اپرون (اگر در پاییندست باشند) جلوگیری کنند. ISها اولین بار در اپرون gal باکتری E. coli که مجموعه سه ژنی و مسئول متابولیسم قسمتی از قند گالاکتوز است، شناسایی شدند.
ترانسپوزون ها برخي اوقات از رپليکوني که در آن الحاق شدهاند خارج میشوند، اما احتمال خروج کمتر از الحاق است. ترانسپوزونها میتوانند حرکت کرده روي مکان ديگري از همان کروموزوم يا کروموزوم متفاوت بنشينند. آنها همچنين میتوانند سازماندهي مجدد کروموزوم را ایجاد کنند.
بررسی فیزیکی DNA الحاقی
برای اثبات وجود DNA الحاقی، یک آزمایش برای بررسی فیزیکی توالی الحاقشونده در نظر گرفتهایم؛ فاژ λ در کنار اپرون gal قرار میگیرد و آن يک راه ساده براي به دست آوردن ذرات فاژ λdgal است که منطقه gal را انتخاب کردهاند. وقتي قطعات IS در gal در داخل λdgal جاي میگیرند، طول کلی برخی از λdgal در کل محلول تغییر میکند، در نتیجه وقتی کل محلول در یک ستون شيب چگالي سزيم کلرايد شناور میشود، گراديان IS فاژهاي λdgal با نرمال قابلمقایسه میگردد (شکل 2). اين مولکولي DNA است که قطعات IS را حمل میکند و بلندتر از DNA نوع معمولی است. آزمايش بهطور واقع اثبات میکند که موتاسيونها بهوسیله درج مقدار عمدهای DNA در داخل اپرون gal ايجاد شدهاند.
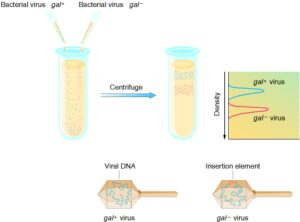
شکل 2) ایجاد جهش توسط DNA الحاقی بهوسیله فاژ λ در اپرون gal، سپس سانتریفیوژ محصول در سدیم کلرید سزیوم، ذرات جهشیافته چگالتر شدهاند. ازآنجاکه ذرات ویروس همه به همان اندازه هستند، افزایش چگالی نشاندهنده مولکول DNA بزرگتر است که با جهش و افزوده شدن قطعهDNA الحاقی بوجود آمده است
تشخيص ISهاي مجزا
آزمایشهای هيبريداسيون نشان میدهد که موتاسيونهاي الحاقی بسيار مختلفي بهوسیله مجموعه کوچکي از سکانسهای الحاقی ايجاد میشوند. بر اساس طرحهای هيبريداسيون متقاطع (Cross Hybridization)، موتانتهاي الحاقی در گروههایی متفاوت قرار داده میشوند. اولين سکانس این گروه با 800 جفت باز در اپرون gal شناسايي شد و IS1 خوانده شد.
دومين سکانس IS2 ناميده شد و طول آن 1350bp است. ما میدانیم که ژنوم استاندارد E. coli غني از قطعات IS است که شامل 8 کپي از IS1 و کپیهای ديگر از انواع IS میشود. بايد تأکید کرد که پیدا شدن ناگهاني سکانسهای الحاقی در هر لوکوس مشخص به اين معنا است که اين عناصر متحرک با قابليت ترانسپوزيشن (جابجایی) در داخل ژنوم هستند. تغییرات تنها وقتي که در داخل يک موقعيت غيرطبيعي قرار میگیرند (مثل يک ژن ساختاري) يا بعضي تغييرات قابلتشخیص در عملکرد، قابل مشاهده هستند.
جهتیابی (گرايش) قطعات IS
قطعات IS در بهمريختگي نسبي، نظم و يا آرايش وقايع مولکولي ژنومي که در آن زندگي میکنند، شرکت میکنند که مهمترین تغییرات ناشی از وجود آنها شامل موارد ذیل است:
1- ترانسپوزيشن: حرکت IS و الحاق آن در مکان ديگر در همان DNA يا مولکول DNA ديگر در سلول
2- غيرفعالسازي الحاقی: الحاق يک IS در داخل يک سکانس کدکننده که باعث از دست رفتن عملکرد آن ژن میشود (Null Mutation).
3- نوترکيبي همولوگ: سبب حذف، وارونگي يا ترکيب مولکولهای DNA میشود.
ترانسپوزونهای پروکاریوتی
در سال 1950 میلادی یک توانایی بسیار جالب و دردسرساز در بیمارستانهای ژاپن در هنگام اپیدمی شیگلوز (اسهال خونی) کشف شد. این بیماری که عامل آن باکتریهای جنس شیگلا هستند در طی این اپیدمی طیف وسیعی از مقاومت با سرعت بالایی نشان میدادند. مطالعات نشان میداد که این باکتریها در ابتدای درمان بهصورت وسیعالطیف مقاوم نبودهاند، اما اکنون به آنتیبیوتیکهایی مثل پنیسیلین، تتراسایکلین، سولفونامید، استرپتومایسین و کلرامفنیکل مقاومت نشان میدهند. محققان ژاپنی متوجه شدند که مقاومت به چندین دارو بهصورت یک بسته واحد ژنتیکی از یک باکتری به باکتری دیگر به ارث میرسد. این مقاومت نسبت به چند آنتیبیوتیک نهتنها میتوانست از یک باکتری به باکتری از همان جنس دیگر منتقل شود، بلکه قادر بود تا بهصورت عرضی جنسها و گونههای دیگر را نیز در برابر آنتیبیوتیکها مقاوم سازد.
این استعداد برای باکتریهای بیماریزا فوقالعاده و برای پزشکان درگیر وحشتناک بود. از دیدگاه متخصصین ژنتیک این موضوع بسیار جذاب و قابلبررسی مینمود. مشاهدات نشان داد که حامل حملکننده این مقاومت از سلول به سلول دیگر حرکت کرده و بهصورت مستقل خود همانندسازی میکند؛ شبیه چیزی که در مورد فاکتورF میدانیم. عوامل این مقاومت بهسرعت از سلول به سلول دیگر انتقال یافته و شبیه فاکتور [9]F در اشریشیا کلی هستند. در واقع هم این عوامل که برای اولین بار کشف شدند، بسیار شبیه فاکتور F بودند. این عناصر در جایگاه پلاسمیدی باکتریها در سیتوپلاسم قرار داشتند و میتوانستند دستههای مختلفی از ژنها را حمل کنند.
عناصر متحرک DNA ضرورتاً انگلهای مولکولي هستند که به ظاهر هيچ عملکردي در زيستایی ارگانیسمهای ميزبان خود ندارند و تنها براي بقاي خود وجود دارند به همين دليل آنها را بهعنوان DNA خودخواه نيز ياد میکنند. انباشت آهستهی اين عناصر در ژنومهاي يوکاريوتي طي زمان تکاملي از ترانسپوزيشن آنها ناشي میشود، همچنين اين عناصر با سرعت بسيار کند بهوسیله حذف قطعات DNA حاوي آنها و يا جمع شدن جهشها ناپديد میشوند به همين دليل عناصر متحرک بسيار آهسته از ژنوم يوکاريوتي حذف میشوند و اکنون به جايگاهي رسیدهاند که قسمت عمده و مهمي از ژنوم بسياري از يوکاريوتها را تشکيل میدهند.
پس يکي از مشخصترین پيامدهاي ترانسپوزيشن، زیانآور بودن آنهاست خصوصاً وقتي که الحاق در داخل ژنهای ضروري اتفاق میافتد. سؤالی که اينجا ايجاد میشود اين است که نگهداري آنها در ژنوم باکتریها در طي تکامل برای چيست؟
اگر عوامل IS اغلب زیانآور بودند از ژنوم باکتري در طولانیمدت حذف میشدند، مگر اینکه بهوسیله ساير عوامل مثل ویروسها يا پلاسميدها بهصورت مکرر با انتقال افقي در ژنوم افزايش پيدا کنند.
صحت يا دقت موتاسيونهاي مفيد وابسته به IS، بسياري از موتاسيونهاي مضر يا طبيعي را متعادل میکند و يکي از دلايل نگهداري بلندمدت آنها در طی تکامل است. نگهداري طولانیمدت IS در سلولها را میتوان بهعنوان يک معامله بين موتاسيونهاي بسيار طبيعي يا مضري که بهآرامی ژنوم باکتري را فلج میکنند و موتاسيونهاي مفيد نادری که به خاطر حضور ISها بهصورت مجاني سوار ژنوم میشوند، در نظر گرفت. سازماندهي مجدد با واسطه IS نقش عمدهای در سازگاري باکتريايي دارند. تثبیت يک موتاسيون مفيد، هم اندازه و هم سرعت موتاسيون را افزايش میدهد.
در قسمتهای بعدی، درباره ساختار فیزیکی، چگونگی عملکرد ترانسپوزون ها و انواع آنها بحث خواهیم کرد.
[1] Transposons
[2] Barbara McClintock
[3] Marcus Rhoades
[4] Transposition
[5] Jumping genes
[6] Roving genes
[7] Mobile genetic elements
[8] Insertion sequences, or insertion-sequence
[9] F factor
https://www.britannica.com/science/transposon
برای دانلود فایل pdf بر روی لینک زیر کلیک کنید
ورود / ثبت نام