مسیر بیوسنتز و ژنتیک آفلاتوکسین B1
عاطفه شجاعیان 1، مژگان سبزعلی زاده 1، تهمینه ابراهیم زاده شیراز 1
1 کارشناسی ارشد میکروبیولوژی، دانشگاه آزاد اسلامی واحد ورامین-پیشوا
به طور کلی آسپرژیلوس-پارازیتیکوس به عنوان يک ساپروفيت روی بقايای گياهی بسياری از گياهان زراعی روی خاک رشد میکند و در سراسر نقاط دنيا يافت میشود و بيشتر تمايل دارد در کشورهايی که اقليم استوايی با ميزان بارش زياد، با دمای بالا و مرطوب دارند، زندگی کند. اين گونه دارای کونيديوفورهایی است بدون ديواره که متمايز از هيف هستند و در بالا به شکل يک کيسه حاوی تعداد زيادی سلول توليد کننده اسپور تخصصی به نام فيالايد در آمدهاند. کلنیهای آ. پارازيتيکوس به رنگ سبز تيره هستند و روی آگار czepak رشد میکنند و با گذشت زمان هم سبز باقی میمانند.
در سنتز آفلاتوکسین چندین گام اصلی در ایجاد کارسینومای هپاتوسلولار القا شده در اثر تماس با آفلاتوکسین B1 مورد تایید واقع شده است که شواهدی مبنی بر آن نشان میدهد که مکانیسم عمل این عامل شامل فعالسازی متابولیک به یک متابولیت آنوتاکسیک، تشکیل ترکیبهای اضافی DNA و مدیفیکاسیون ژن TP53 میباشد. حضور همزمان هپاتیت B میزان وقوع تومورهای هپاتیک در انسان را افزایش میدهد. آفلاتوکسین B1 متداولترین و بالقوهترین آفلاتوکسین میباشد. این ماده به طور عمده در کبد متابولیزه و تبدیل به آفلاتوکسین 9و8 اگزواپوکسید آفلاتوکسین B1 میشود که این ماده ثانویه یک گوانین AFB1-NT پیش موتاژن را ایجاد میکند که در نهایت منجر به موتاسیونهایی از نوع ترانسورژن G به T میگردد. در سرطان هپاتوسلولار انسانی در مناطقی که تماس با آفلاتوکسین بسیار است بیش از 50% تومورها شامل یک موتاسیون خاص AGG به AGT در کدون 249 ژن مهارکننده تومور TP53 میباشند.
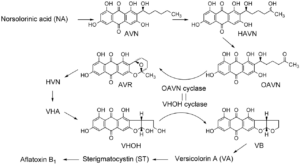
آفلاتوکسينها گروهی متشکل از متابوليتهای ثانويه مشتق از پلیکتايدها هستند که عمدتا توسط گونههای آسپرژيلوس فلاووس و آسپرژيلوس پارازيتيکوس توليد میشوند. آنها سمی، موتاژن و کارسينوژنيک برای گونههای مختلف حيوانات و انسانها هستند. از اين رو آلودگی محصولات کشاورزی با اين سموم، تاثيرات بسيار مضری برای سلامتی حيوانات و انسانها به همراه دارد. آلودگی با آفلاتوکسين در محصولاتی از قبيل گردو، فندق، بادامزمينی، ذرت و پنبه رخ میدهد. قارچهای مولد آفلاتوکسين محصولات را در مرحله قبل از چيدن و هم بعد از چيدن میتوانند آلوده نمايند .مسير بيوسنتز آفلاتوکسين در تلاش برای فهم بهتر کنترل مولکولی بيوسنتز آفلاتوکسين، به صورت گستردهای مطالعه شده است. حداقل ١٨ واکنش آنزيمی در بيوسنتز آفلاتوکسين شرکت دارند که با سنتز هگزانوآت (hexanoate) از استات (acetate) آغاز میشود. در مراحل اوليه بيوسنتز آفلاتوکسين، نورسولورینیک اسید (norsolorinic acid {NA}) به آورانتین (averantin{AVN}) توسط آنزیم ردوکتازی که nor-1 کد میکند، تبدیل میشود. آنزیم مونواکسیژنازی که توسط avnA کد میشود، تبدیل AVN را به اتم 5-هیدروکسی آورانتین (HAVN) کاتالیز میکند.HAVN یک پیشساز آفلاتوکسینهاست.HAVN در حضور NAD یا NADP به آوروفین(averufin، AVR) تبدیل میشود و این واکنش NAD را به جای NADP برای کوآنزیم ترجیح می دهد. chang و همکاران گزارش دادند که تخریب ژن adhA که در گروه ژنی آفلاتوکسین آسپرژیلوس پارازیتیکوس قرار دارد، باعث تجمع HAVN میگردد که نشان دهنده کد کردن آنزیم HAVN دهیدروژناز توسط این ژن است. HAVN توسط HAVNدهیدروژناز به OAVN تبدیل میشود که OAVN برای فعالیتش به NAD نیاز دارد. سپس OAVN توسط آنزیم OAVN سیکلاز به AVR تبدیل میشود.McCormick و همکارانش گزارش کردند که آوروفانین (averufanin) به AFB1 تبدیل میشود که نشان میداد آوروفانین در مرحله بین AVN و AVR نقش دارد. برخلاف آن Sakuno و همکارانش نشان دادند که HAVN یک پیشساز آفلاتوکسین است و HAVN به AVR تبدیل میشود نه آوروفانین. همچنین ثابت کردند که HAVN یک ماده ناپایدار است که به راحتی از طریق دهیدراسیون خود به خودی به آوروفانین تبدیل میشود.
مسیر HAVN به OAVN از دو واکنش تشکیل شده است:
1-واکنش دهیدروژناسیون از HAVN به OAVN توسط آنزیم adhA دهیدروژناز
2-واکنش دهیدراسیون (سیکلیزاسیون،cyclization) از OAVN به AVR توسط آنزیم OAVN سیکلاز.
به طور کلی آفلاتوکسین B1 از سنتز اسید چرب (FAS) و نیز سنتز پلیکتاید(pks) که هر دو به عنوان مسیر سنتز نورسولورونیک اسید شناخته شدهاند به دست میآید. بیوسنتز آن با سنتز هگزانوآت توسط FAS آغاز میشود که این ماده یک واحد آغاز کننده برای pks نوع I تکرار شونده میباشد. pks، هفت دنباله مانوئیل COA به هگزانوآت اضافه میکند تا ترکیب c2o پلیکتاید را تشکیل دهد. سپس یک ردوکتاز(E1) کاهش کتون روی زنجیره جانبی نورسولورونیک اسید را کاتالیز میکند تا averantin تشکیل شود. سپس این ماده توسط دو آنزیم هیدروکسیلاز (E2) و الکل دهیدروژناز (E3) به averufin تبدیل میشود. در نهایت averufin توسط P450 اکسیداز (E4) اکسید میشود. بدین ترتیب حلقههای اتری باز شده و Versiconal استات تشکیل میشود. آنزیم E5 هیدرولیز استیل را کاتالیز کرده و الکل اولیه را در Versiconal تشکیل میدهد. بعد از تاثیر آنزیمهای E6 و E7 ، VersicolorinB به dihydrobis furan تبدیل میشود. آنزیمهای E8 و E9 ، VersicolorinA را به دیمتیل استریگماتوسیستئین تبدیل میکند، سپس S-آدنوزیل متیونین (SAM) دو گروه هیدروکسیل روی بخش xanthone دیمتیل استریگماتوسیستئین توسط آنزیمهای E10 و E11 متیله میشوند و در مرحله آخر آنزیم E12 حلقه آروماتیک را شکسته شده و یک کربن مفقود میشود و آخرین اتفاق برای تشکیل آفلاتوکسین B1 رخ میدهد.
شکل 1: مسیر سنتز آفلاتوکسین B1
پس از جذب این ماده توسط گروهی از آنزیمهای سایتوکروم، P450 در کبد متابولیز میشود و بسته به استعداد ژنتیکی گونه تبدیل به محصولات متابولیک مثل آفلاتوکسیول، آفلاتوکسین Q1 ، آفلاتوکسین P1 و آفلاتوکسین M1 میشود. علاوه بر متابولیتهای ذکر شده، آفلاتوکسین epoxide 8.9 نیز تشکیل میشود. مقدار مشخص از این متابولیتها قادر است القای جهش در DNA را سبب شود.
تولید آفلاتوکسین تحت تاثیر شرایط رشد (منبع کربن و نیتروژن و PH محیط) و چرخه سلولی میباشد. اثرات منبع کربن، نیتروژن و PH همگی به ترتیب مرتبط با فعالیت عمومی فاکتورهای رونویسی pac c، Arc A ،cre A میباشد. هر سه این فاکتورها تولید آفلاتوکسین را به واسطه کنترل سطح بیان aflR، aflJ، Pac c تنظیم میکنند. مسیرهای سیگنالینگ دقیق این فاکتورهای رونویسی هنوز به طور دقیق شناخته نشده است. همچنین عوامل تغذیهای و محیطی بر بیوسنتز آفلاتوکسین اثرگذار است. جایگاههای اتصال Pac c و Arc A در ناحیه بینژنی aflR-aflJ شواهد محکمی مبنی بر این است که بیان ژن توسط سیگنالهای محیطی تنظیم میشود. در مطالعات مشاهده شده است که آسپرژیلوسها نسبت به گونههای دیگر به طرق مختلف به نیترات پاسخ میدهند و این تفاوت میتواند به علت اختلاف در تعداد جایگاههای GATA مجاور به جایگاه شروع aflJ باشند. ژنهای دیگر در بیوسنتز خوشه آلفا شامل جایگاههای اتصال Pac c و Arc A در موقعیتهای مهم در پروموتور خود هستند که میتواند بر بیان ژن آنها تاثیر گذارد. به طور کلی بیان ژنهای نیترات ردوکتاز و نیتریت ردوکتاز مستلزم مهار متابولیت نیتروژن و القا توسط نیترات است. لازم به ذکر است که تمام ژنهای مسیر بیوسنتز آفلاتوکسین فاقد جایگاه cre A در پروموتور خود هستند، بنابراین در معرض مهار کاتابولیتی کربن که با واسطه فاکتور رونویسی cre A است قرار نمیگیرند. فاکتور رونویسی دیگری که به قند پاسخ می دهد Rgt1 میباشد که برای تنظیم بیان مولکول ترانسپورتر گلوکز ضروری است. جایگاه احتمالی Rgt1 در ناحیه پروموتور آسپرژیلوس پارازیتیکوس است که میتواند در تنظیم بیان آن موثر باشد. راه دیگری که بر بیان ژن آفلاتوکسین اثر میگذارد شاید القای سیگنالینگ وابسته به G1-protein باشد. در سلولهای آسپرژیلوس، سیگنالینگ پروتئین G رشد قارچ و تشکیل آفلاتوکسین را تنظیم میکند.
تمام ژنهایی که تا به حال نشان داده شده که در مسیر بیوسنتز آفلاتوکسین دخالت دارند در یک ناحیه kp 70 از ژنوم آسپرژیلوس پارازیتیکوس واقع شدهاند. این خوشههای ژنی یکسان متشکل از 24 ژن میباشند که با کاتالیز یک یا چند مرحلهای، مستقیما در بیوسنتز آفلاتوکسین نقش دارند.
از لحاظ ژنتیکی، تولید آفلاتوکسین هم در سطح رونویسی و هم در سطح پس از رونویسی تنظیم میشود. مطالعات نشان میدهد که یک فاکتور رونویسی اختصاصی به نام Afl R در آسپرژیلوس پارازیتیکوس روی خوشه ژنی آفلاتوکسین اثر میگذارد. آنالیزهای بیوانفورماتیکی نشان داده است که Afl R به خانواده خوشه دو هستهای تعلق دارد که به صورت هومودایمر به طور اختصاصی به DNA هدف خود متصل میشود. Afl R یک فاکتور رونویسی با عملکرد مثبت است.
متابولیسم آفلاتوکسین B1 به خوبی در انسان و حیوانات آزمایشگاهی مطالعه شده است.
CYP1A2/2B6/3A4/3A5/3A7/GSTM1 از جمله آنزیمهایی هستند که در متابولیسم آفلاتوکسین در انسانها دخالت دارند. میزان دخالت این آنزیمها در متابولیسم آفلاتوکسین B1 در محیط زنده نه تنها به میل اتصالی آنها بستگی ندارد بلکه به میزان بیان آنها در کبد انسان نیز ارتباط ندارد. آنزیم CYP3A4 در تشکیل اگزواپوکسید و آفلاتوکسین Q1 دخالت دارد، در حالی که CYP1A2 میتواند مقداری اگزواپوکسید، مقدار زیادی اندواپوکسید و آفلاتوکسین M1 تولید کند. آفلاتوکسینهای M1/Q1 به ترتیب توسط CYP1A2 و 3A4 تولید شدهاند و در ادرار افراد قرارگرفته در معرض آفلاتوکسین وجود دارند. در انسانها همانند گونههای دیگر اتصال DNA و سرطانزایی آفلاتوکسین B1 ناشی از تبدیل آن به آفلاتوکسین B1 اگزواپوکسید 9 و 8 توسط CYP3A4 میباشد. این اپوکسید بسیار واکنشی و فعال بوده و اصلیترین واسطه آسیب سلولی میباشد. CYP3A5 برخلاف CYP3A4 آفلاتوکسین را به طور عمده به 9 و 8 اگزواپوکسید متابولیزه میکند اما در کاتالیز 3-هیدروکسیلاسیون آفلاتوکسین B1 به متابولیت آفلاتوکسین 1Q حدود 100برابر ضعیفتر عمل میکند. بیان آنزیمهای درگیر از متابولیسمهای آفلاتوکسین را میتوان با استفاده از عوامل مهار شیمیایی تعدیل کرد، به طوریکه از تشکیل DNA-adduct و هپاتوکارسینوژنز جلوگیری شود.Olltipraz یک ماده مهارکننده شیمیایی است که میزان کنژوگاسیون گلوتاتیون را افزایش داده و از فعالیت برخی از آنزیم های سیتوکروم p450 جلوگیری میکند. آفلاتوکسینها و ویروسهای کبدی عوامل مهمی در افزایش میزان وقوع فوقالعاده بالای کارسینومای سلول کبدی میباشند. آلودگی با HBV ممکن است متابولیسم آفلاتوکسین را افزایش دهد. قابل توجه است که فعالیت گلوتاتیون دترانسفراز در کبد انسان در حضور عفونت HBV کاهش مییابد، در موشهای ترانس ژنتیک HBV آسیب کبد با افزایش بیان آنزیم های سیتوکروم P450 مرتبط است.
افلاتوکسین B1 روی برخی از رسپتورهای هستهای تاثیرگذار است. اثر آفلاتوکسین روی رسپتورهای PXR، CAR، ALR و سایتوکرومهای هدف آنها یعنی P450 روی هپاتوسیتهای انسان میباشد. اثر آفلاتوکسین روی رسپتورهای مذکور باعث افزایش بیان سیتوکروم P2B6، CYP3A5 میشود. مطالعات نشان داده که اثر آفلاتوکسین روی این رسپتورها موجب فعال شدن آنها میشود. در مطالعاتی نشان داده شد که آفلاتوکسین B1 بیان mRNA ژنهای PXR، CAR، CYP2A5، CYP3A5 و CYP2C9 را افزایش داده است.
CAR و AHR: با اتصال به لیگاند منجر به فعالسازی رسپتور میشود که در این صورت رسپتور متعهد میشود انتقال هستهای را انجام دهد.
PXR: در غیاب لیگاند به صورت PXR-PXR یعنی متصل به هترودایمر اختصاصی خود میباشد که در این صورت این حالت به صورت متراکم میشود.کروماتین بر مهار بیان ژن اثر میگذارد اما در صورت وجود لیگاند مربوطه، رسپتور ACTIVATOR را به کار گرفته و مهارکننده های مهاجر آزاد میشود. این فرآیند منجر به افزایش میزان رونویسی از ژنهای هدف میشود.
https://medlabnews.ir/%d9%85%d8%a7%db%8c%da%a9%d9%88%d8%aa%d9%88%da%a9%d8%b3%db%8c%d9%86%e2%80%8c%d9%87%d8%a7%db%8c-%d8%a2%d8%b3%d9%be%d8%b1%da%98%db%8c%d9%84%d9%88%d8%b3-%d9%be%d8%a7%d8%b1%d8%a7%d8%b2%db%8c%d8%aa%db%8c/
برای دانلود فایل pdf بر روی لینک زیر کلیک کنید
ورود / ثبت نام