تولید پلاستیک زیستی (پلیهیدروکسی آلکانوات) بهوسیله میکروارگانیسمها
(قسمت دوم)
وهاب پیرانفر (کارشناش ارشد)، محمد عرفانی (کارشناس ارشد)، دکتر رضا میرنژاد (دانشیار دانشگاه)
در قسمت قبل در خصوص پلاستیک زیستی (پلیهیدروکسی آلکانوات) ازجمله ساختار، ویژگیها و خصوصیات فیزیکی مطالبی ارائه شد که در این بخش به راههای مختلف سنتز این ماده اشاره میگردد.
سنتز پلیهیدروکسی آلکانوات
ژنها و آنزیمهای دخیل در فرآیند
در این فرآیند سه آنزیم که توسط سه ژن کد میشوند، مشارکت دارند. ژن phaA آنزیم بتاکتوتیولاز[1] را کد میکند. این آنزیم باعث تبدیل دو مولکول استیل کوآنزیم آ[2] به فرم استواستیل کوآنزیم آ[3] میشود. در مرحله بعد احیاء استواستیل کوآنزیم آ به (R)-3-هیدروکسی بوتیریل کوآنزیم آ[4] بهوسیله آنزیم استواستیل کوآنزیم آ ردوکتاز[5] صورت میگیرد. این آنزیم که یک آنزیم وابسته به NADPH میباشد بهوسیله ژن phaB کد میشود. واکنش نهایی، پلیمریزاسیون مونومرهای (R)-3-هیدروکسی بوتیرات کوآنزیم آ میباشد که بهوسیله آنزیم پلیهیدروکسی آلکانوات سنتئاز[6] کاتالیز میشود که این آنزیم توسط ژن phaC کد میشود.
پلیهیدروکسی آلکانوات سنتئاز در باکتری رالستونیا اتروفا[7] با طیف اندکی از سوبستراها واکنش میدهد؛ سوبستراهایی با طول زنجیره ۳ تا ۵ کربنه مناسب واکنش میباشند، ازاینرو پلیهیدروکسی آلکانواتهای حاصله حاوی مونومرهای زنجیره کوتاه میباشند. تمامی این سه آنزیم در سیتوزول سلول باکتری قرار دارند. تغییر در نوع و مقدار نسبی منابع کربن در محیط رشد باکتری میتواند در ایجاد پلیهیدروکسی آلکانواتهایی با مونومرهای متفاوت نقش داشته باشد، به طور مثال اضافه کردن پروپیونیک اسید[8] یا والریک اسید[9] در محیط گلوکز، تولید را به سمت ایجاد کوپلیمرهای مرکب از هیدروکسی بوتیرات و هیدروکسی والرات هدایت میکند. البته در این مسیر چگالش پروپیونیل کوآنزیم آ[10] به استیل کوآنزیم آ با واسطه یک کتوتیولاز مجزا انجام میشود (3- کتوتیولاز توسط ژن bktB کد میشود). احیاء 3-کتو والریل کوآنزیم آ به (R)-3-هیدروکسی والریل کوآنزیم آ و پلیمریزاسیون که متعاقباً سبب تشکیل پلیهیدروکسی بوتیرات کوهیدروکسی والرات میشود بهواسطه آنزیمهایی که در سنتز پلی هیدروکسی آلکانوات نقش دارند، به نامهای استواستیل کوآنزیم آ ردوکتاز و پلیهیدروکسی آلکانوات سنتئاز کاتالیز میشود.
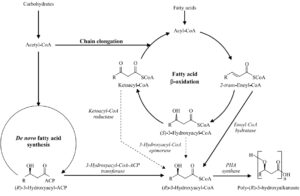
شکل (1): مسیر متابولیکی سنتز پلی هیدروکسی بوتیرات
تمام پلیهیدروکسی آلکانوات سنتئازهایی که تاکنون شناخته شده است را میتوان در چهار کلاس با توجه به فاکتورهای اختصاصی سوبسترا و زیرواحدهای تشکیلدهنده، تقسیمبندی نمود. ژنهای کدکننده آنزیمهای کلیدی سنتز پلیهیدروکسی آلکانوات بهوسیله چند میکروارگانیسم تولیدکننده زیستبسپار، کلون و بیان گردیده است.
اولین ژن phaA بود که با استفاده از آنتیبادیهای ضد تیولاز[11] از باکتری زاگلوا رامیگرا[12] بدست آمد. بعدها مشخص شد که ژن phaB در این گونهها و پاراکوکسی دنیتریفیکانس[13] و رایزوبیوم ملیلوتی[14] در اپرونهای مشابهی وجود دارد، درحالیکه ژن phaC در اپرون متفاوتی قرار دارد.
در رالستونیا اتروفا، گونههای اسینتوباکتر[15]، آلکانیژنز لاتوس[16] و سودوموناس اسیدوفیلا[17] ژنهای pha در اپرون phaABC قرار دارد، اگرچه توالی این سه ژن در همه این گونهها یکسان نیست و در برخی موارد ژنوم این گونهها بیش از یک کپی از اپرون را حمل میکنند.
در تعدادی از گونهها از قبیل کروماتیوم وینسوم[18]، تایوسیتس ویولاس[19] آ، تایوکاپسا فنیجی[20] و ساینکوسیتیس گونه PCC 6803[21] پلیهیدروکسی آلکانوات سنتئاز به طور عمده سنتز پلیهیدروکسی آلکانواتهای زنجیره کوتاه را کاتالیز میکند. در این میان ژنهای دیگری نیز هستند که با کد کردن برخی آنزیمها به طور غیرمستقیم در سنتز پلیهیدروکسی آلکانوات نقش دارند. ژن پلیهیدروکسی آلکانوات سنتئاز در ائروموناس کاویا[22] در مجاورت ژن phaJ قرار میگیرد. phaJ آنزیم انول کوآنزیم آ هیدراتاز[23] را کد میکند، این آنزیم با هیدراتاسیون 2-انول کوآنزیم آ، مونومر (R)-3-هیدروکسی آسیل کوآنزیم آ را برای سنتز پلیهیدروکسی آلکانوات از طریق بتااکسیداسیون اسید چرب تأمین میکند.
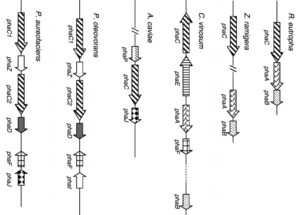
شکل (2): موقعیت ژنهای سنتزکننده پلیهیدروکسی آلکانوات در باکتریهای مختلف
برخلاف رالستونیا اتروفا، باکتریهای بورخولدریا کاریوفیلی[24]، سودوموناس اولئوورانس[25] و سودوموناس آئروژینوزا[26] قادر به ساخت پلیهیدروکسی آلکانواتهایی با زنجیره متوسط هستند. اپرون phaC1ZC2D در این ارگانیسمها حاوی دو ژن phaC میباشد که بهوسیله ژن phaZ از هم جدا شدهاند. ژن phaC1 و phaC2 دو پلیمراز از خانواده آلفا/بتا هیدرولاز[27] را کد میکنند. ژن phaZ یک پلیهیدروکسی آلکانوات دپلیمراز را کد میکند. نقش phaD هنوز ناشناخته است و به نظر میرسد پروتئین حاصله از این ژن در تشکیل پلیهیدروکسی آلکانوات موردنیاز میباشد.
پلیهیدروکسی آلکانوات در باکتری سودوموناس اولئوورانس میتواند پلیمریزاسیون طیف وسیعی از مونومرها را کاتالیز کند که نتیجه آن تولید پلیمرهایی با وزن مولکولی بسیار بالا و خواص الاستیکی بهتر میباشد. چند میکروارگانیسم دیگر خوشه ژنی phaFI که در فرودست اپرون phaC1ZC2D قرار دارد را حمل میکنند. phaI در فرمدهی و تثبیت گرانولها دخیل است، درحالیکه phaF هم نقش تنظیمی دارد و هم در تثبیت گرانولها نقش دارد. در غیاب پیشماده پلیهیدروکسی آلکانوات، phaC1ZC2D و phaI بیان نمیشوند. زمانی که مونومرهای زنجیره متوسط تولید میشوند، پروتئین PhaF از DNA خارج میشود و پلیهیدروکسی آلکانوات اولیه ساخته میشود. تحت این شرایط پروتئینهای PhaF و PhaI با زنجیره پلیمری در حال توسعه هیدروفوبیک برهمکنش میدهند و در فرمدهی گرانول مشارکت میکنند. آخرین ژن phaP میباشد که پروتئینی با وزن مولکولی بسیار پایین به نام فازین[28] را کد میکند؛ این پروتئین با اتصال گرانولها به یکدیگر سبب افزایش تولید پلیهیدروکسی بوتیرات میشود.
بتااکسیداسیون[29] اسید چرب
کاتابولیسم اسیدهای چرب یکی از رایجترین مسیرها برای تأمین مونومرهای هیدروکسی آلکانوات در سنتز پلیهیدروکسی آلکانوات میباشد. حدواسطهای ساخته شده با تجزیه آلکانوئیک اسید یا اسیدهای چرب بهوسیله بتااکسیداسیون میتوانند سوبسترای هیدروکسی آلکانوئیل را برای ساخت پلیهیدروکسی آلکانواتهای زنجیره متوسط ایجاد کنند. این مسیر در چندین باکتری از قبیل سودوموناس اولئوورانس و سودوموناس فراجی[30] که قادر به تولید پلیهیدروکسی آلکانوات زنجیره متوسط از اسید آلکانوئیک یا اسید چرب هستند، مشاهده شده است. ترکیب مونومری پلیهیدروکسی آلکانواتهای تولید شده به طور مستقیم با سوبسترای استفاده شده برای رشد باکتری در ارتباط است و معمولاً دارای مونومرهایی با دو اتم کربن کمتر نسبت به اتمهای کربن سوبسترا هستند. هرچند تیپ وحشی اشریشیاکلی نمیتواند پلیهیدروکسی آلکانوات را ذخیره کند، اما اشریشیاکلی دستورزی شده با استفاده از مسیر متابولیکی اسیدهای چرب میتواند پیشمادههای هیدروکسی آلکانوات کوآنزیم آ را برای ذخیرهسازی پلیهیدروکسی آلکانوات تأمین نماید.
حدواسطهای حاصله از مسیر بتااکسیداسیون اسید چرب شامل انول کوآنزیم آ، 3-کتوآسیل کوآنزیم آ و (S)-3-هیدروکسی آسیل کوآنزیم آ میباشند که این حد واسطها میتوانند بهعنوان پیشسازهای (R)-3-هیدروکسی آسیل کوآنزیم آ که مستقیماً در سنتز پلیهیدروکسی آلکانواتهای زنجیره متوسط شرکت دارند، مورداستفاده قرار بگیرند. با این حساب سنتز پلیهیدروکسی آلکانواتهای زنجیره متوسط در سلول اشریشیاکلیهایی امکانپذیر است که در مسیر بتااکسیداسیون تغییر پیدا کردهاند، در این وضعیت ژن پلیهیدروکسی آلکانوات سنتئاز باکتری سودوموناس در اشریشیاکلی ناقص قرار میگیرد و با بیان مشترک دیگر ژنها باعث افزایش تبدیل حدواسطهای بتااکسیداسیون به (R)-3-هیدروکسی آسیل کوآنزیم آ میشود، با توجه به اینکه R-3-هیدروکسی آسیل کوآنزیم آ پیشساز ضروری سنتز پلیهیدروکسی آلکانواتهای زنجیره متوسط میباشد.
مثال این ژنها که بیان مشترک دارند با پلیهیدروکسی آلکانوات سنتئاز برای افزایش تولید عبارتند از paaF,paaG,ydbU در اشریشیاکلی، phaJ در سودوموناس آئروژینوزا، سودوموناس پوتیدا و ائروموناس کاویا، maoC و yfcX در اشریشیاکلی، fabG(در سودوموناس آئروژینوزا و اشریشیاکلی و rhlG در سودوموناس آئروژینوزا.
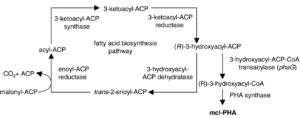
شکل (3): بیوسنتز de novo اسید چرب
بیان مشترک دیگر ژنهای مسیر بتااکسیداسیون میتواند به طور قابلتوجهی تولید پلیهیدروکسی آلکانوات را در اشریشیاکلی نوترکیب افزایش دهد، برای مثال آنزیم آسیل کوآنزیم آ دهیدروژناز در اشریشیاکلی بهوسیله ژنهای yafH و fadE کد میشود. این آنزیم هیدراتاسیون آسیل کوآنزیم آ را به انول کوآنزیم آ، کاتالیز میکند. بیان مشترک yafH به همراه phaC و phaJ باعث افزایش مخزن انول کوآنزیم آ میشود. با افزایش مخزن پیشمادهها، اشریشیاکلی نوترکیب ۴ برابر بیشتر پلیهیدروکسی آلکانوات زنجیره کوتاه و متوسط و کوپلیمر پلی (3-هیدروکسی بوتیرات-کو-3-هیدروکسی هگزانوات) را نسبت به سلولهایی که تنها ژنهای phaC و phaJ در آنها بیان شده بود، ذخیره میکنند. همینطور بیان ژن yafH در اشریشیاکلی سبب افزایش مقدار هیدروکسی هگزانوات در کوپلیمر پلی(3-هیدروکسی بوتیرات-کو-3-هیدروکسی هگزانوات) تولید شده توسط ائروموناس هیدروفیلا[31] میشود.
در برخی باکتریها پلیهیدروکسی آلکانوات بهواسطه بیوسنتز de novo اسید چرب انجام میگیرد (شکل 3). برخلاف سودوموناس اولئوورانس و سودوموناس فراجی (که حدواسطهای بتا اکسیداسیون را استفاده میکنند)، باکتریهای سودوموناس آئروژینوزا و سودوموناس پوتیدا هنگامی که بر روی سوبستراهای غیرمرتبط مثل گلوکز رشد میکنند، سبب تولید پلیهیدروکسی آلکانواتهای زنجیره متوسط میشوند. این به سبب استفاده سودوموناس اولئوورانس و سودوموناس فراجی از اسید چرب بهعنوان منبع کربن در مسیر بتااکسیداسیون میباشد که طی آن 3-هیدروکسی آسیل کوآنزیم آ (سوبسترای پلیهیدروکسی آلکانوات سنتئاز) تولید میشود.
از طرف دیگر بیوسنتز اسید چرب در سودوموناس آئروژینوزا و سودوموناس پوتیدا راه اصلی سنتز 3-هیدروکسی آسیل کوآنزیم آ میباشد. این باکتریها در طی رشد بر روی منبع کربن سبب سوختوساز موادی مانند کربوهیدرات، استات و اتانول به استیل کوآنزیم آ میشوند. ژن phaG آنزیمی مرتبط با سنتز پلیهیدروکسی آلکانوات و بیوسنتز de novo اسید چرب را کد میکند. محصول این ژن، (R)-3-هیدروکسی آسیل-ACP (حد واسط بیوسنتز اسید چرب) را به مشتقات کوآنزیم آ تبدیل میکند، بنابراین بیان phaG در اشریشیاکلی، قابلیت جدیدی را برای ساخت پیشمادههای اسید چرب زنجیره متوسط ایجاد میکند و بیان آن در باکتریهای سودوموناس اولئوورانس و سودوموناس فراجی باعث تولید پلیهیدروکسی آلکانواتهای زنجیره متوسط از منابع کربنی غیرمرتبط با 3-هیدروکسی دکانوئیک اسید[32] از قبیل گلوکز و فروکتوز میشود. تنها زمانی phaG به همراه phaC قادر به ساخت پلیهیدروکسی آلکانوات زنجیره متوسط در اشریشیاکلی است که بیوسنتز de novo اسید چرب آنها بهوسیله تریکلوسان[33] محدود شده باشد.
[1] β-ketothiolase
[2] Acetyl-CoA
[3] Acetoacetyl-CoA
[4] (R)-3-hydroxybutyryl-CoA
[5] Acetoacetyl-CoA reductase
[6] PHA synthase
[7] Ralstonia eutropha
[8] Propionic Acid
[9] Valeric Acid
[10] Propionyl-CoA
[11] Anti-thiolase
[12] Zoogloea ramigera
[13] Paracoccus denitrificans
[14] Rhizobium meliloti
[15] Acinetobacter
[16] Alcaligenes latus
[17] Pseudomonas acidophila
[18] Chromatium vinosum
[19] Thiocystis violacea
[20] Thiocapsa pfennigii
[21] Synechocystis spp.
[22] Aeromonas caviae
[23] Enoyl-CoA hydratase
[24] Burkholderia caryophylli
[25] Pseudomonas oleovorans
[26] Pseudomonas aeruginosa
[27] α/β hydrolase
[28] Phasin
[29] β-oxidation
[30] Pseudomonas fragi
[31] Aeromonas hydrophila
[32] 3-Hydroxydecanoicacid
[33] Triclosan
تولید پلاستیک زیستی (پلیهیدروکسی آلکانوات) بهوسیله میکروارگانیسمها (1)
برای دانلود فایل pdf بر روی لینک زیر کلیک کنید
ورود / ثبت نام