
نقش ترانسپوزونها در پروکاریوتها
(قسمت دوم)
وهاب پیرانفر (کارشناس ارشد)، محمد عرفانی (کارشناس ارشد)، دکتر رضا میرنژاد (دانشیار دانشگاه)
ساختار فیزیکی ترانسپوزونها
اگر DNA يک پلاسميد که داراي مقاومت دارويي است (مثلاً ژنهای مقاومت به کانامايسين را حمل میکند)، دناتوره گردد تا يک تک رشتهاي شکل بگيرد و سپس رناتوره شود، بعضي از رشتهها يک شکل غيرمعمول در زير ميکروسکوپ الکتروني ايجاد ميکنندکه شبیه ساختار آب نبات چوبي میباشد (شکل 1). این ساختار، يک DNA دو رشتهاي میباشد که از طريق قرار گرفتن دو سکانس IR (Inverted repeat) در پلاسميد ايجاد میگردد (شکل 2). مطالعات نشان داده که سکانسهای IR در اکثر موارد يک جفت هستند. ژنهای مقاومت دارويي يا ساير تواناییهای ژنتيکي که بهوسیله پلاسميد حمل میشوند اين سکانسهای IR در سر آبنبات چوبي قرار دارند. سکانسهای IR با ژنهایی که همراهشان است مجموعاً يک ترانسپوزون ناميده میشوند. ترانسپوزونهاي بلندتر از IS المنتها هستند، زيرا ژنهایی که کدکننده پروتئين هم هستند، باقيمانده پلاسميد ژنهای انتقالدهنده مقاومت (Resistance Transfer Function) را حمل میکنند که منطقه RTF ناميده میشوند (شکل 3).
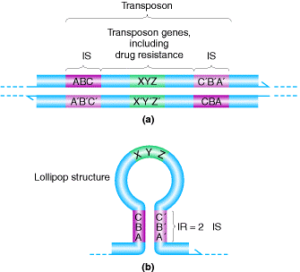
شکل 1: نمایی در سطح نوکلئوتیدها برای ساختار آبنبات. این ساختار ترانسپوزون نامیده میشود (رنگ بنفش). بخش a، ترانسپوزون را قبل از دناتوره شدن نشان میدهد.
نکته: به توالی معکوس در قسمت موردنظر توجه داشته باشید. شکل ب شکل آبنبات را نشان میدهد که بهآرامی رشته از حالت دناتوره به رناتوره تغییر شکل میدهد و ساختار معکوس به یکدیگر میچسبند. ژنهای ترانسپوزون در بین این دو رشته معکوس قرار میگیرند.

شکل 2: دو ترانسپوزون را نشان میدهد که دارای توالی IR (تکراری معکوس) متفاوت میباشند و حامل ژنهای مقاومت در برابر آنتیبیوتیک هستند. (a): Tn9 که منطقه IR کوتاهتری دارد چراکه دو عنصر IS در یک جهت هستند. (b): Tn10 یک منطقه بزرگ تکراری معکوس به خاطر دو مؤلفه در جهت مخالف
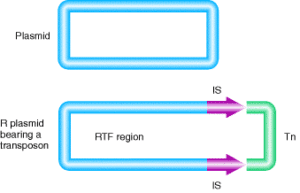
شکل 3) یک ترانسپوزون داخل یک پلاسمید. RTF نشاندهــــــنده عملکرد resistance-transfer از پلاسمید است. ترانسپوزون شامل عناصر الحاقی و ژن مقاومت به آنتیبیوتیک است.
ژنهای تنظیمکننده جابجایی ترانسپوزونها
ژن مربوط به مقاومت نسبت به آنتیبیوتیکها در واقع عابری است که توسط DNA ترانسپوزون حمل میشود. ترانسپوزونها دارای دو خصوصیت مهم هستند؛ یکی اینکه 40-20 نوکلئوتید موجود در یکی از انتهاهای آزاد بهطور معکوس در انتهای دیگر قرار دارد (جهت دو توالی انتهایی در خلاف یکدیگر است و به آنها توالی تکراری معکوس گفته میشود) و دیگر اینکه بسیاری از ترانسپوزونها (احتمالاً همه آنها) آنزیم ترانسپوزاز را کد مینمایند. این آنزیم دخول ترانسپوزون به جایگاههای جدید را به عهده دارد.
گفته میشود که کم شدن غلظت آنزیم ترانسپوزاز سبب کم شدن انتقالها در سلول میگردد. بهطور متوسط در هر تقسیم سلولی کمتر از یک مولکول ترانسپوزاز بهوسیله ترانسپوزون Tn10 ساخته میشود که این مقدار تنها برای انجام یک ترانسپوزیشن (انتقال) در ده میلیون تولیدمثل کافی است. چنانچه به کمک مهندسی ژنتیک پلاسمیدی ساخته شود که ترانسپوزاز بیشتری تولید کند ملاحظه میگردد که عمل ترانسپوزیشن Tn10 هزار بار بیشتر میشود. به نظر میرسد که سرعت عمل ترانسپوزیشن بهطور طبیعی از طریق تغییر میزان تولید ترانسپوزاز کنترل میگردد.
اخیــــــراً عاملی که بیان و عمل ترانسپوزاز را کنترل میکند کشف شــــــــــــده است. پروموتر Tn10 و بعضی دیـــــــــــــــــگر از ترانسپوزونها حاوی GATC اســــــــت که در کلیباسیلها گروه آدنیـــــــــــــــــن ایـــــــــــــــن توالی توســـــــــــــط آنزیم ســـــــــــلولی دَم متــــــیلاز متیله شده است. ازآنجاکه توالی
5´GAT3´
نیز در جهت ´5 به ´3 همان GATC است هر دو رشته در یک جایگاه متیله میشوند و در این حالت پروموتر غیرفعال خواهد بود. هنگامی که DNA همانندسازی میکند یک دقیقه یا بیشتر وقت لازم است که آدنین توالی GATC تازه سنتزشده، توسط دَم متیلاز متیله شود. در طی این مدت کوتاه که جایگاه فوق بهصورت نیمه متیله است پروموتر فعالیت بیشتری نشان میدهد و در این حالت بلافاصله بعد از همانندسازی DNA مقدار کمی ترانسپوزاز ساخته میشود.
از طرف دیگر جایگاه موجود در DNAی Tn10 که آنزیم ترانسپوزاز بر روی آن عمل میکند نیز دارای توالی GATC است، بنابراین در طول مدتی که تنها یک رشته پروموتر متیله شده است، جایگاه فوق سوبسترای بهتری برای ترانسپوزاز میباشد. بدین ترتیب ملاحظه میشود که Tn10 درست بعد از همانندسازی تمایل به ترانسپوزیشن دارد. هنوز شرایط دیگر سلولی که احتمالاً بر روی متیلاسیون DNA تأثیر میگذارد و در اثر آن سرعت ترانسپوزیشن تغییر میکند و یا راههای دیگر تنظیم ترانسپوزیشن شناخته نشده است. بعضی ترانسپوزونها آنزیم دیگری به نام رزولواز را کد میکنند که در مرحله دوم ترانس پوزیشن مؤثر هستند.
در مورد Tn3 آنزیم رزولواز فعالیت ثانویه مستقل دیگری دارد، بدین ترتیب که سبب سد کردن بیان ژن خود و ژن ترانسپوزاز میشود و از طریق بیان آنها را در حد طبیعی پایین نگه میدارد.
چگونگی حرکت ترانسپوزونها
بررسی توالی DNA ترانسپوزونهای باکتریها مانند Tn3 و نیز بررسی جایگاه اتصال آنها به DNA هدف تا حدی مکانیسم ترانسپوزیشن را روشن کرده است؛ اولاً حرکت ترانسپوزونها دقیق است و سبب حمل کلیه توالیهای دخولی جایگاه قدیمی میشوند، درحالیکه بر توالیهای مجاور اثری نمیگذارند. این موضوع تنها زمانی میتواند صادق باشد که آنزیمی (بهاحتمال قوی ترانسپوزاز) انتهاهای ترانسپوزون را تشخیص دهد ضمناً انتهاهای فوق باید یکسان باشند. چون آنها جایگاههای اتصال مشترک ترانسپوزاز هستند. ثانیاً در جایگاه دخول، 3 الی 12 باز از DNA هدف (تعداد نوکلئوتیدها بستگی به نوع ترانسپوزون دارد) مضاعف میشوند و یک کپی از آنها در هر یک از انتهای ترانسپوزون باقی میماند، بنابراین لازمه مضاعف شدن جایگاه هدف در هنگام ترانسپوزیشن، سنتز DNA است و این اختلاف مهمی با نوترکیبی در جایگاه ویژه که هنگام انتگره شدن فاژ لامبدا بوجود میآمد، دارد. ثالثاً با آنکه اکثر ترانسپوزونها عمدتاً به هر ناحیهای از ژنوم جدید میروند، ولی حرکت آنها تصادفی نیست بلکه به توالیهای DNA معینی حمله مینمایند. بعضی از این ترانسپوزنها روی توالیهای متقارن چهار تا شش جفت بازی (درست مشابه آنچه سوبسترای آنزیمهای محدود کننده است) اثر میگذارند.
مطالعات انجامشده بر روی آنزیمهایی مانند انتگراز فاژ لامبدا و توپوایزومراز که سبب قطع و اتصال مجدد DNA میشوند، میتوانند بهعنوان مدلی جهت توضیح چگونگی عمل ترانسپوزاز باشند. احتمالاً ابتدا ترانسپوزاز به انتهای ترانسپوزون و نیز توالی DNA هدفی که ترانسپوزون در آن دخول خواهد یافت، متصل میشود و سپس برشهایی با انتهای چسبنده مشابه آنچه آنزیمهای محدودکننده ایجاد میکردند در DNA هدف و ترانسپوزون ایجاد میکند و سپس انتهاهای آزادشده ترانسپوزون به جایگاه هدف متصل میشوند و بدین طریق دو مولکول DNA به هم ملحق میگردند (ترانسپوزون بین دو انتهای چسبنده DNA اولیه قرار میگیرد). بههرحال برحسب نوع ترانسپوزون یا شرایط خاص رشد، دو نوع اتصال ممکن است انجام شود؛ در ترانسپوزیشن ساده در انتهاهای ترانسپوزون برشهای اضافه بوجود میآید بهطوریکه تنها ترانسپوزون کامل وارد DNA جدید میگردد و این امر سبب باقی ماندن شکافی در DNA میزبان قدیمی میشود که کشنده بوده و ممکن است سبب مرگ آن گردد. بههرحال DNA جدید که حالا واجد ترانسپوزون شده است با قرار گرفتن چند نوکلئوتید (این عمل توسط DNA پلیمراز صورت میگیرد) ترمیم شده، تمامیت خود را بدست میآورد (شکل 4).
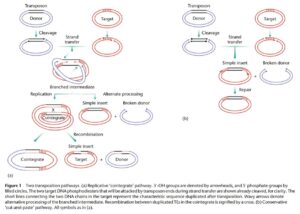
شکل 4: دو راه انتقال ترانسپوزونها
بنابراین ترانسپوزیشن ساده عمل حفاظتی است که در آن ترانسپوزون وارد یک جایگاه جدید میشود. نوع دوم ترانسپوزیشن همانندسازیکننده است که در آن ترانسپوزون مضاعف میشود و تنها یک کپی بهسوی DNA هدف میرود و کپی دوم باقی میماند. در این حالت DNA مبدأ (میزبان) تغییر نمیکند و برشهای ثانویهای در آن ایجاد نمیشود و با اتصال کل DNA میزبان و هدف به یکدیگر و انجام همانندسازی (جهت پر کردن شکاف) دو مولکول بهطور کامل به یکدیگر ملحق میشوند. در مرحله بعد عمل رزولواز در جایگاهی معین، برش ایجاد میگردد. بدین ترتیب دو قطعه DNA مبدأ و هدف از یکدیگر جدا میشوند و با تکمیل عمل همانندسازی شکافها یک بار دیگر پر میگردند و در این حالت DNA مبدأ از بین نمیرود و یک نسخه کامل از آن به DNA هدف میرسد. درحالیکه چنانچه هر یک از این مراحل تغییر کند انواع مختلف فعالیتهای ترانسپوزونها ظاهر میشوند، مثلاً اگر آنزیم رزولواز وجود نداشته باشد DNA مرکبی که شامل DNA مبدأ و مقصد است بوجود میآید (مانند پلاسمید R1).
ضمناً امکان دارد در محل ایجاد برش، سکانس مولکولهای DNA مرکب با یکدیگر همخوانی نداشته، در این صورت دو مولکول DNA بازآرایی شده مستقل پدید خواهد آمد. با بررسی دقیقتر چنانچه ترانسپوزیشن در یک مولکول واحد DNA رخ دهد (DNAهای مبدأ و مقصد یکی باشند) حذف یا معکوس شدن DNA صورت میگیرد و سبب تغییر شدید ساختمان کروموزوم میشود.
با توجه به آنکه عمل ترانسپوزیشن باکتریوفاژ MU که هم یک فاژ است و هم یک ترانسپوزون، در عصاره سلولی هم انجام میگردد، میتوان انتظار داشت که به کمک آن بتوان دقیقاً چگونگی حرکت ترانسپوزونها را درک نمود.
تجارب اخیر نشان میدهند که حرکت بعضی از ترانسپوزونهای جانداران عالیتر خصوصاً آنهایی که مربوط به رتروویروسها هستند با ترانسپوزونهای باکتریهایی مانند Tn3 کاملاً متفاوت است؛ مثلاً در مخمر، ابتدا از روی عناصر Ty (TY element) رونویسی انجام میگردد و سپس به کمک آنزیم رونویسیکننده معکوس از روی آن DNA ساخته میشود و در اثر نوترکیبی، این قطعه DNA، از طریق مکانیسمی که هنوز بهخوبی شناخته نشده است در کروموزوم میزبان جای میگیرد.
در قسمت بعدی درباره انواع ترانسپوزونها بحث خواهیم کرد.
نقش ترانسپوزونها در پروکاریوتها (1)
برای دانلود پی دی اف بر روی لینک زیر کلیک کنید
ورود / ثبت نام